 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库解码木薯块根3类蛋白修饰图谱,解锁淀粉积累与抗逆调控密码

英文标题: Deciphering the Atlas of Protein Acetylation, 2‑Hydroxyisobutyrylation, and Malonylation in Developing Cassava Roots
中文标题: 解析木薯块根发育过程中蛋白质乙酰化、2-羟基异丁酰化和丙二酰化修饰图谱
发表期刊: Journal of Agricultural and Food Chemistry
影响因子: 6.2
研究背景
蛋白质翻译后修饰(Posttranslational modifications, PTMs)是调节蛋白质功能、影响植物生长发育和环境胁迫响应的关键机制。其中,赖氨酸乙酰化(Kac)、2-羟基异丁酰化(Khib)和丙二酰化(Kma)是近年来发现的三种新型赖氨酸酰化修饰,它们在多种生物过程中发挥着重要作用。这些PTMs通过改变酶活性、亚细胞定位、蛋白质稳定性和蛋白质-蛋白质相互作用来精细调控蛋白质功能。
木薯(Manihot esculenta)作为全球8亿多人口的主食,是一种重要的淀粉块根作物。其块根的发育,特别是淀粉的积累,对其产量至关重要。尽管已有大量组学研究关注木薯根系发育、采后生理和逆境胁迫,但针对Kac、Khib和Kma等PTMs在木薯中调控蔗糖/淀粉代谢和淀粉积累的全面研究仍是空白。本研究首次构建木薯发育中块根中Kac、Khib和Kma修饰的全局蛋白质组学图谱,旨在揭示这些PTMs在木薯根系发育和生物/非生物胁迫响应中的调控作用。通过深入分析这些修饰的特征、共存模式及其在关键代谢途径中的功能富集,为阐明PTM驱动的植物调控分子机制提供了新的见解。
研究结论
01. 木薯块根中Kac、Khib和Kma的全局分析
研究以发育中的木薯块根为实验材料,通过免疫印迹与质谱分析,明确了Kac、Khib、Kma为木薯中独立且广泛存在的蛋白修饰系统(图1A),并且完成了三类修饰的全局鉴定(图1B-D)。在修饰位点与蛋白分布上,Khib修饰位点数量最多,且其蛋白中高修饰位点数占比显著高于Kac和Kma,三类修饰蛋白均以少于5个修饰位点为主(图1E-G);修饰频率方面,三类蛋白均以低频率修饰为主,Khib蛋白出现高频率修饰的数量最多(图1H-J);对于亚细胞定位,三类修饰蛋白均主要分布在叶绿体、细胞质和细胞核,而同时含三种修饰的蛋白则以细胞质定位为主(图1K-N)。
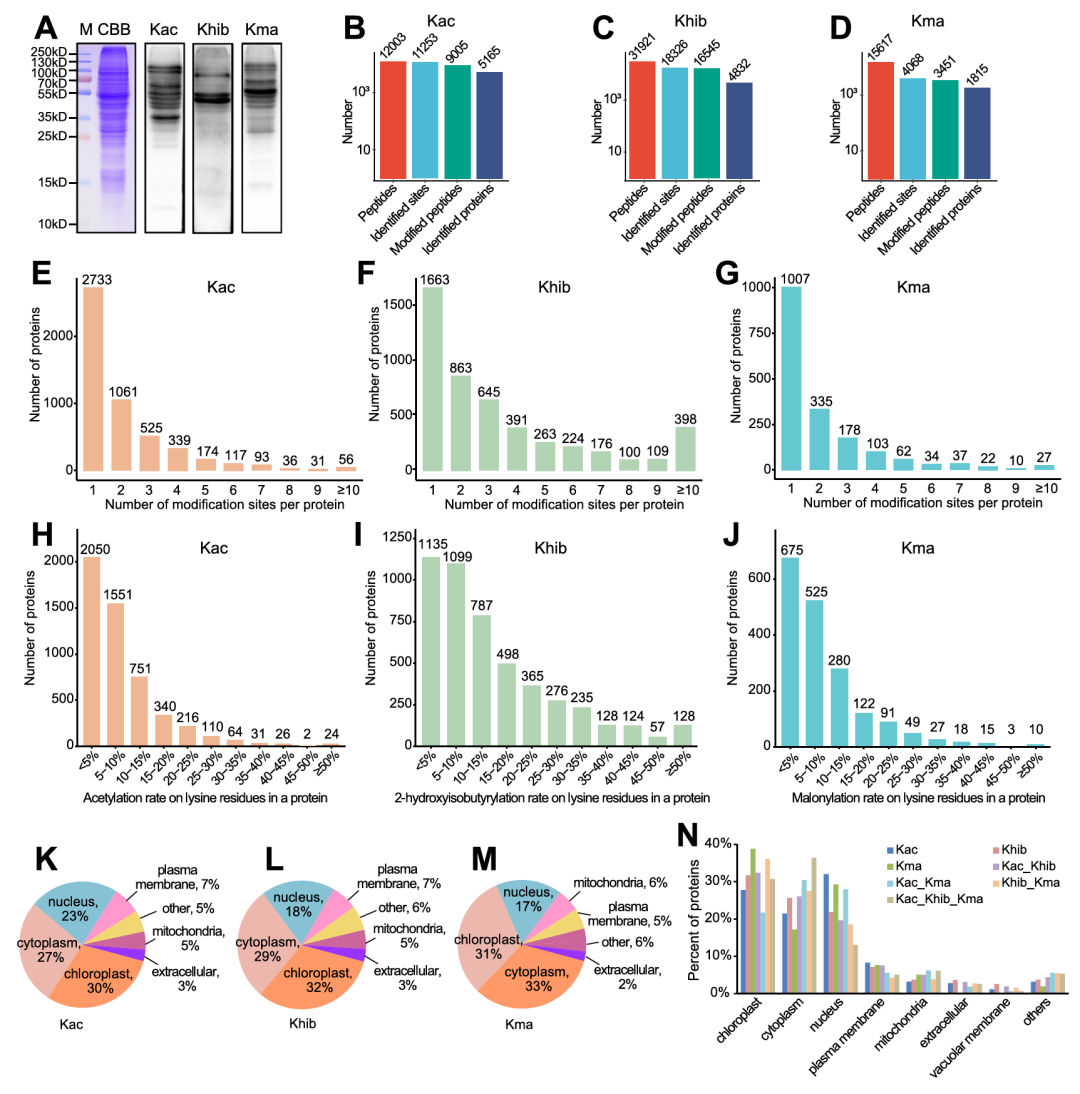
图1. 木薯块根中Kac、Khib和Kma的全局鉴定与特征分析
02.Kac、Khib和Kma修饰的特征分析
为明确三类修饰的氨基酸偏好特征,研究进一步解析了木薯中Kac、Khib、Kma修饰位点的氨基酸序列特征(图2A)与基序规律(图2B)。整体而言,三类修饰位点侧翼均富集丙氨酸(A)、赖氨酸(K)、缬氨酸(V)和(Y)、组氨酸(H)存在位点特异性富集:G在Kac/Kma上下游均富集、仅在Khib下游富集,C仅富集于Kac/Kma近端,Y和H为Kac位点特有。基序鉴定发现,Kac的高表达基序数量最多,且16个基序富集倍数超5.0,Khib和Kma则极少;Kac基序多在+1位富集L、I等残基,超60%的Khib基序侧翼含K残基,Kma无高富集倍数基序。这些结果为木薯蛋白修饰位点的预测提供了关键依据。
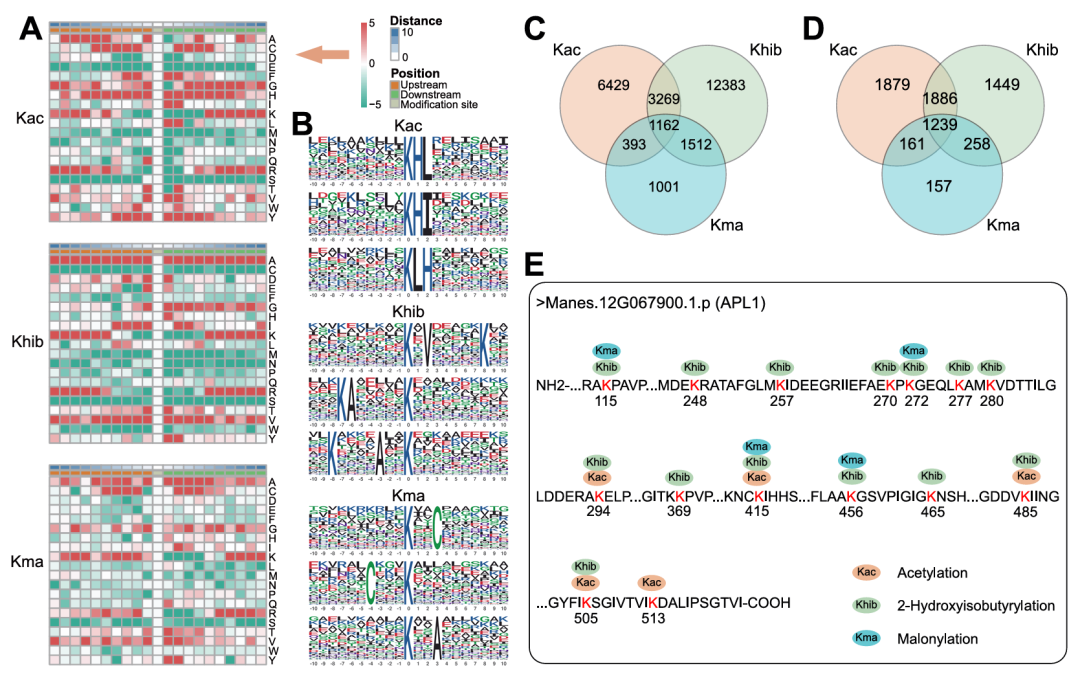
图2. Kac、Khib和Kma修饰位点的特征分析及修饰蛋白的重叠性研究
03.Kac、Khib和Kma修饰的重叠分析
基于上述修饰特征,研究还分析了Kac、Khib和Kma修饰的重叠性(图2C-E),以明确这些修饰是否会共发生在同一赖氨酸位点或同一蛋白上。结果显示,1239个蛋白、1162个位点同时存在三类修饰,Kac与Khib的共修饰蛋白数(1886个)远高于Kac-Kma、Khib-Kma组合;单一修饰分析发现,仅Kac、Khib修饰的蛋白数远多于Kma,表明前两者的调控范围更广。其中,淀粉积累关键蛋白APL1为三类修饰共现的典型,其含有5个Kac、14个Khib、4个Kma位点,且K415为三类修饰的共同位点(图2E)。由此可见,木薯中单个赖氨酸残基可以经历多种PTMs,这可能使得蛋白质功能能够进行协调调控。
04.组蛋白上Kac、Khib和Kma位点的保守性
组蛋白修饰在表观遗传调控和基因表达调控中具有关键作用。通过解析木薯组蛋白上Kac、Khib、Kma修饰的分布特征(图3A-B),总计鉴定出23个带119个修饰位点的组蛋白,其中H2B、H3、H4可被三种修饰共同修饰,H2A仅存在Kac修饰,且无组蛋白同时携带Kac和Kma修饰。此外,不同组蛋白的特定位点存在固定的共修饰模式,如H2B的K24、K61为三类修饰共定位点(图3C)。在序列保守性方面(图3C),H2A和H4的多个Kac位点、H4的三个Khib位点在木薯、甘蔗、水稻中高度保守;相反,H2B的Kac和Khib修饰保守性极低,木薯与甘蔗间无共享位点;尽管木薯与水稻的H3部分Kac位点也存在保守性,但因缺乏数据,所以暂未发现保守Kma位点。综合而言,这些保守修饰位点表明Kac和Khib在木薯与其他植物间存在进化保守性,可能参与木薯的重要生物学功能调控。
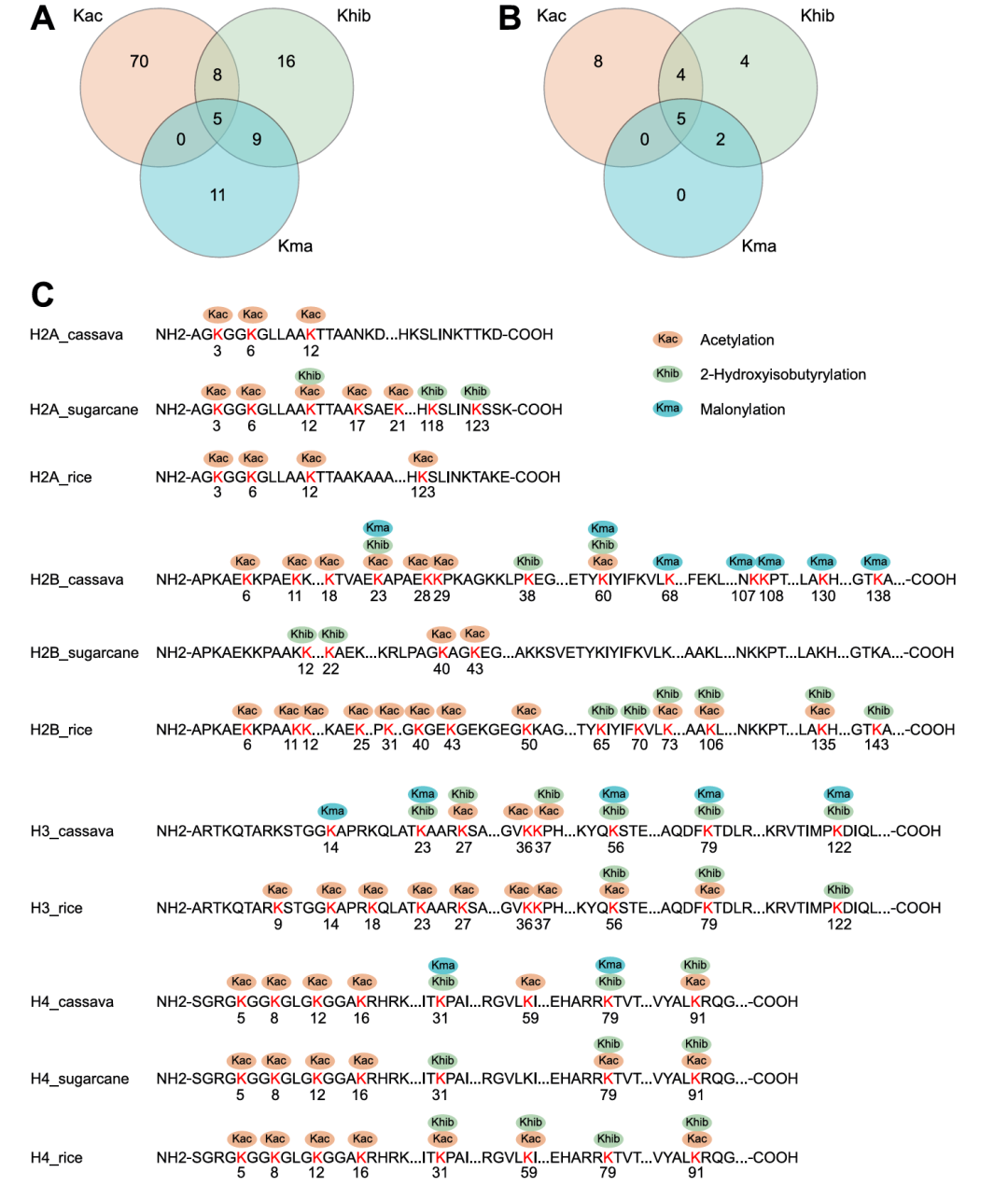
图3. Kac、Khib和Kla在组蛋白上的修饰
05.Kac、Khib和Kma蛋白的功能富集分析
借由COG/KOG功能注释、GO富集及KEGG富集分析,Kac、Khib和Kma修饰蛋白在非组蛋白中的生物学功能被进一步阐明。COG/KOG注释结果显示,Kac、Khib和Kma修饰蛋白参与多种生物学过程,主要包括碳水化合物代谢、能量产生与转化、翻译后修饰、信号转导、翻译以及RNA加工与修饰(图4A)。GO富集分析表明,这些修饰蛋白参与肽转运、二羧酸代谢、三羧酸循环代谢过程和丙酮酸代谢(图4B)。KEGG富集分析发现,Kac、Khib和Kma修饰蛋白显著富集于三羧酸循环、丙酮酸代谢、糖酵解/糖异生、果糖和甘露糖代谢以及戊糖磷酸途径(图4C)。
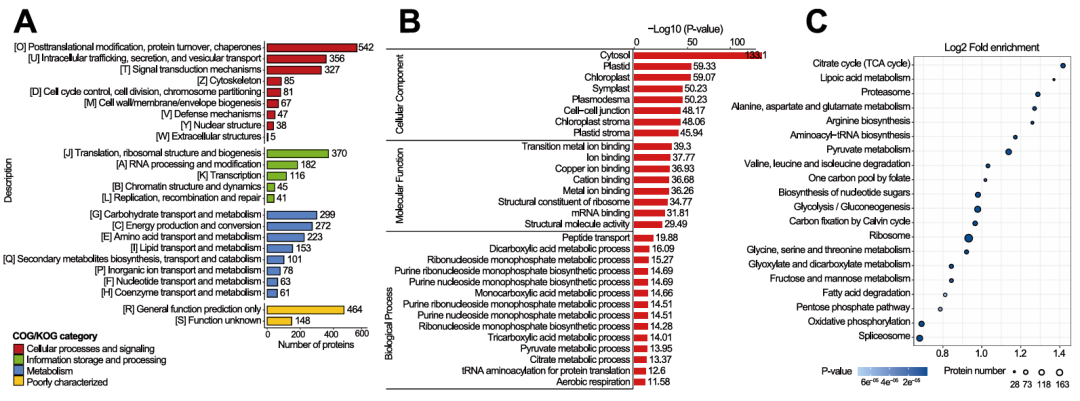
图4. Khib蛋白的功能富集分析(Kac和Kma蛋白详见图S1-S2)
06.Kac、Khib和Kma蛋白参与蔗糖和淀粉代谢
蔗糖和淀粉代谢对木薯块根的生长发育及淀粉积累至关重要。聚焦Kac、Khib、Kma修饰对木薯蔗糖和淀粉代谢的调控作用,研究总计鉴定出29个相关修饰蛋白,覆盖蔗糖降解、淀粉合成与降解全流程(图5)。其中,11个核心蛋白(5个蔗糖降解、3个淀粉合成、3个淀粉降解相关)同时发生三类修饰,13个蛋白受Kac和Khib协同修饰,仅3个蛋白为单一修饰。结果而言,Kac与Khib在该代谢通路中协同调控作用显著,三类修饰共同构成了广泛且精密的调控网络,为木薯块根淀粉积累提供了重要的翻译后修饰层面调控基础。
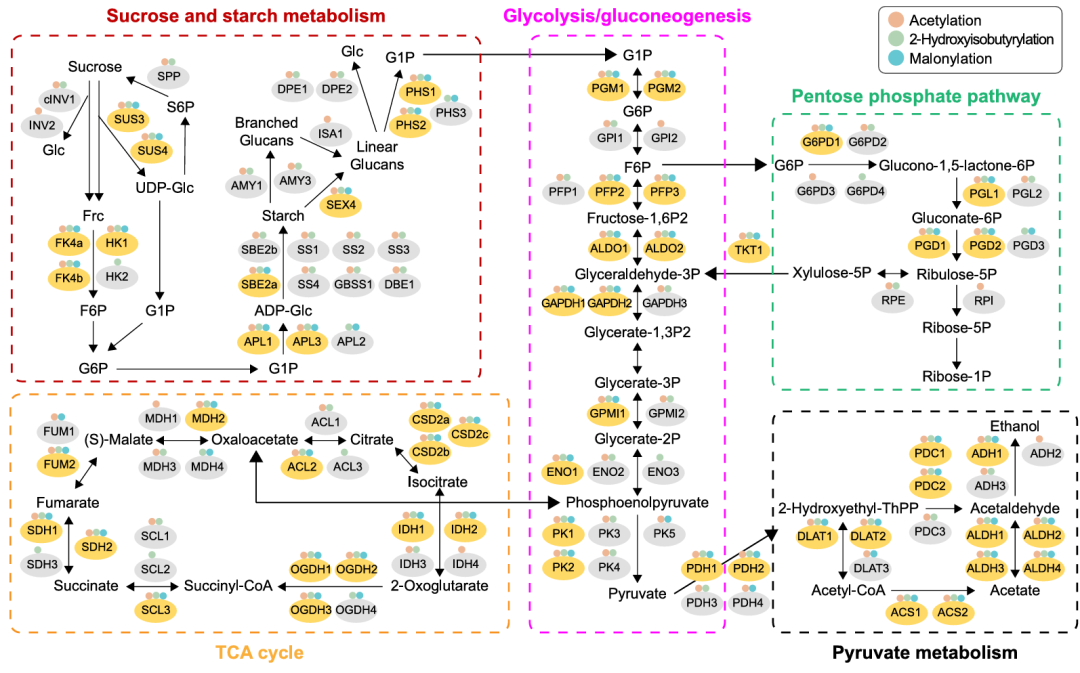
图5. Kac-、Khib-和Kla-修饰蛋白在能量代谢中发挥作用
07.Kac、Khib和Kma蛋白参与糖酵解/糖异生、磷酸戊糖途径、TCA循环和丙酮酸代谢
聚焦核心碳代谢通路(糖酵解/糖异生、戊糖磷酸途径、TCA循环及丙酮酸代谢),Kac、Khib、Kma相关的修饰特征也被逐一揭示(图5)。结果显示,在四类通路中分别有22、12、26、19个相关蛋白被修饰,且核心特征高度一致:均有大量蛋白(依次为12、5、14、13个)同时发生三类修饰,覆盖通路全程;多数剩余蛋白以Kac和Khib共同修饰为主,单一修饰或Kac+Kma、Khib+Kma组合的蛋白占比极少。简言之,Kac、Khib和Kma在木薯能量代谢的调节中起着关键作用,并且大多数蛋白质至少受到其中两种PTM的修饰。
08.Kac、Khib和Kma蛋白参与木质素生物合成
木质素是植物细胞壁的关键结构成分,为植物生长发育提供机械强度,Kac、Khib、Kma也广泛参与着该通路的调控(图6A-B)。具体而言,总计23个修饰蛋白被鉴定,覆盖全合成流程,其修饰类型呈现多样化特征:7个核心蛋白同时发生三类修饰,6个蛋白受Kac和Khib协同修饰,双修饰组合还包括Kac+Kma、Khib+Kma;单一修饰以Kac为主(6个蛋白),仅1个蛋白为Khib单一修饰,未发现Kma单一修饰的蛋白。可见,三类修饰通过多模式组合调控木质素生物合成的关键节点,为木薯细胞壁结构形成和机械强度维持提供了重要的翻译后修饰调控支撑。
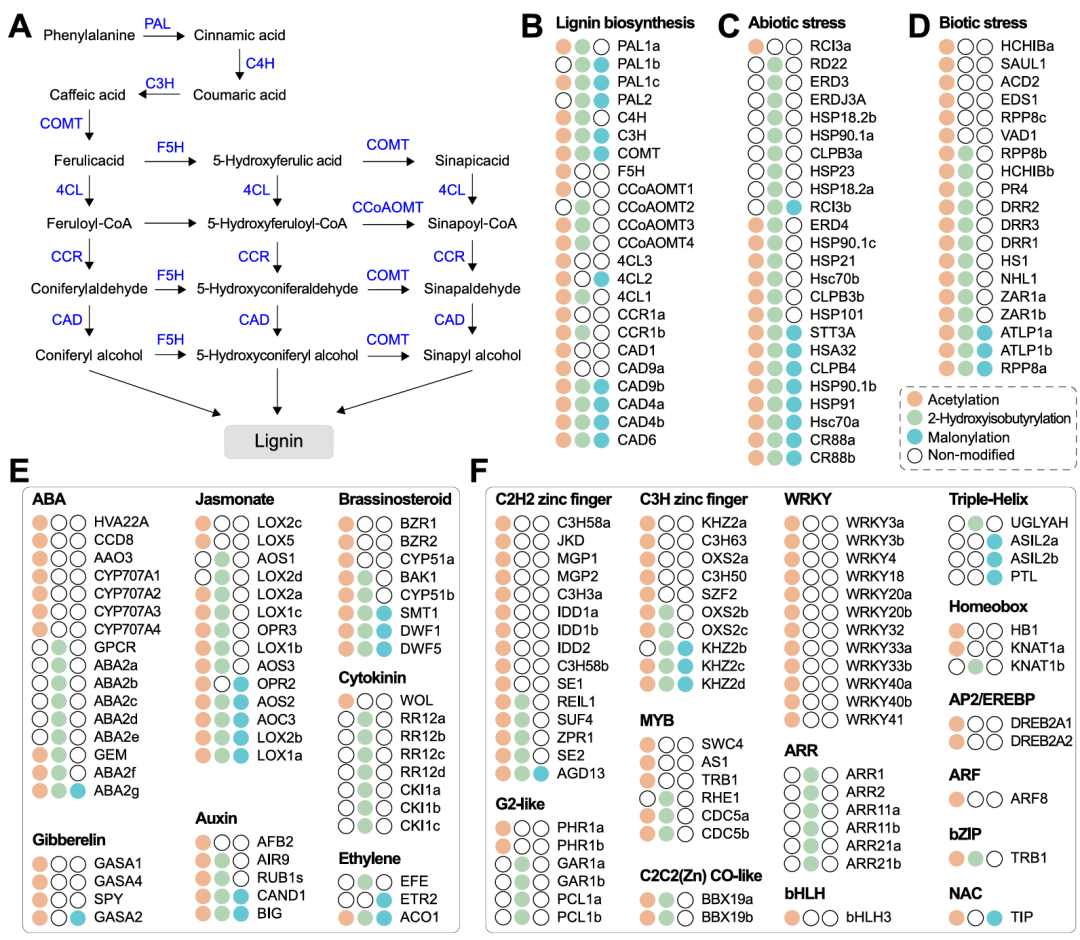
图6. Kac-、Khib-和Kla-修饰蛋白在木质素生物合成、非生物和生物胁迫、激素代谢以及转录因子中发挥作用
09.Kac、Khib和Kma蛋白参与胁迫代谢、激素调控及转录因子功能
对于生物和非生物胁迫,总计43个修饰蛋白被鉴定(图6C-D)。其中,3个生物胁迫蛋白(ATLP1a、ATLP1b、RPP8a)和8个非生物胁迫蛋白(STT3A、HSA32等)同时发生三类修饰;17个蛋白为双修饰类型,含有16个Kac与Khib组合。值得注意的是,96%的非生物胁迫蛋白存在Khib修饰,而所有生物胁迫蛋白均有Kac修饰,表明两类胁迫通路中PTM修饰偏好存在显著差异。
对于激素代谢相关蛋白,木薯中共有58个被修饰(图6E),以脱落酸(ABA,16个)、茉莉酸(14个)、油菜素内酯(8个)和细胞分裂素(8个)相关蛋白居多。ABA合成(ABA2g)、茉莉酸合成(AOS2等)等通路的部分蛋白同时发生三类修饰;茉莉酸和生长素相关蛋白多为Kac与Khib双修饰,赤霉素相关蛋白多仅含Kac修饰,细胞分裂素相关蛋白多仅含Khib修饰,ABA相关蛋白则半数含Kac、半数含Khib修饰,不同激素通路的PTM偏好性不同。
此外,70个转录因子(TF)相关蛋白被修饰(图6F),以C2H2锌指(15个)、WRKY(12个)和C3H锌指(10个)家族为主。仅3个TF蛋白(AGD13等)同时发生三类修饰,且不同家族存在修饰偏好:WRKY家族仅含Kac修饰,ARR家族仅含Khib修饰,Triple-Helix家族多为Kma修饰,C2H2锌指等家族以Kac修饰为主,近半数同时含Khib修饰。
综上所述,Kac、Khib和Kma修饰的蛋白质参与调节胁迫代谢、激素和转录因子,并在其子类别或子途径中表现出不同的修饰偏好。
研究总结
本研究首次构建了木薯块根发育过程中Kac、Khib和Kma的全局蛋白质修饰图谱,共鉴定出11253个Kac位点、18326个Khib位点和4068个Kma位点,分别分布于5165、4832和1815个蛋白中,明确了三类修饰为木薯中独立且广泛存在的调控系统,其修饰蛋白主要定位于叶绿体、细胞质和细胞核,且存在大量共修饰现象。通过特征分析揭示了三类修饰位点的氨基酸偏好与特异性基序,物种比对发现组蛋白上部分Kac和Khib位点具有进化保守性。此外,功能研究表明,修饰蛋白广泛参与蔗糖/淀粉代谢、糖酵解/糖异生等核心碳代谢通路,以及木质素合成、胁迫响应、激素调控和转录因子功能调控,且不同通路中存在显著的修饰偏好性,证实Kac、Khib和Kma通过协同或特异性修饰,在木薯块根发育和生物/非生物胁迫适应中发挥关键调控作用,为解析植物翻译后修饰介导的分子机制及木薯遗传改良提供了重要理论依据与资源支持。





