 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库Developmental constraints mediate the summer solstice reversal of climate effects on the autumn phenology of European beech
Abstract
Accurate projections of temperate tree growing seasons under climate change require representing developmental constraints that determine tree resource allocation. Recent work has identified the summer solstice (21 June) as a phenological “switch point” with pre-solstice warming advancing autumn phenology and post-solstice warming delaying it. However, the flexibility of this switch point remains poorly understood. We performed trans-solstice climate manipulation experiments on potted European beech (
Fagus sylvatica
) saplings to test (i) how spring leaf-out timing and June-August temperatures influence end-of-season timing (bud set and leaf senescence [50% loss of leaf chlorophyll content]), and (ii) whether daytime and nighttime temperatures before and after the solstice have different effects, given that trees primarily grow at night. Bud set and leaf senescence responses were tightly coupled (
R
2
= 0.49), with bud responses being generally stronger. Each day delay in spring leaf-out delayed bud set by 0.24 ± 0.06 days and senescence by 0.22 ± 0.08 days. Post-solstice full-day cooling in July delayed autumn phenology in late-leafing individuals (bud set +4.9 ± 2.6 days; senescence +3.1 ± 2.8 days) but had negligible impact on early-leafing trees (bud set +1.4 ± 2.6 days; leaf senescence +2.2 ± 2.8 days). Conversely, August full-day cooling advanced both stages. Daytime cooling before the solstice had no effect, while after the solstice it advanced autumn phenology. Nighttime cooling always delayed bud set. These findings support the Solstice-as-Phenology-Switch model and highlight the central role of developmental progression in constraining growing seasons. Faster early-season development –especially under nighttime warming– moves trees past the switch earlier, increasing sensitivity to late-season cooling and thereby triggering earlier autumn phenology. To improve growing season length projections, phenology models should account for these developmentally-mediated and diel-specific temperature responses.
Introduction
Climate shifts are leading to rapid, species-specific changes in phenology and ecosystem productivity (
Boisvenue & Running, 2006
;
Menzel
et al
., 2006
;
Yang & Rudolf, 2010
;
Thackeray
et al
., 2016
). In temperate forests, changes in the timing of spring leaf-out, autumn leaf-senescence and bud set are modifying water, energy and carbon cycles (
Peñuelas
et al
., 2009
;
Richardson
et al
., 2013
), with extended growing seasons increasing net ecosystem carbon uptake by up to 9.8 gC m
−2
day
−1
(
Keenan
et al
., 2014
). Therefore, understanding the interaction between climate change and temperate forest phenology is pivotal to improving forecasts of community dynamics and carbon sequestration.
The past few decades have seen delays in the onset of temperate autumn phenology, but these changes are much smaller in magnitude compared to the advances in spring leaf-out observed during the same period (
Gill
et al
., 2015
;
Piao
et al
., 2019
). This is unexpected, given that experiments have demonstrated a high sensitivity of leaf senescence to autumn warming, with phenological responses even surpassing the temperature sensitivity (days per °C) of spring leaf-out (
Fu
et al
., 2018
). One possible explanation for this discrepancy is that other factors may counterbalance the effects of autumn warming, with some studies finding that earlier leaf-out leads to earlier leaf senescence (
Fu
et al
., 2014
;
Keenan & Richardson, 2015
;
Zani
et al
., 2020
). This connection between spring and autumn phenophases could be due to developmental and nutrient constraints that affect carbon source-sink dynamics in temperate trees (
Paul & Foyer, 2001
;
Zani
et al
., 2020
;
Zohner
et al
., 2023
;
Gessler & Zweifel, 2024
).
The Solstice-as-Phenology-Switch hypothesis, supported by experiments and monitoring data, posits that warmer air temperatures before the summer solstice accelerate tree growth and development, leading to an earlier onset of leaf senescence, whereas warmer temperatures after the solstice slow the progression of leaf senescence (
Zohner
et al
., 2023
). The opposite is equally true in colder years, with pre-solstice cooling delaying autumn phenology and post-solstice cooling advancing it. This reversal in temperature effects provides an explanation for the less pronounced shifts in autumn phenology compared to spring (
Piao
et al
., 2019
). The effect reversal after the solstice points to the role of photoperiod in regulating plant physiology (
Bauerle
et al
., 2012
;
Petterle
et al
., 2013
;
Singh
et al
., 2017
), with trees becoming increasingly responsive to cooling temperatures as days continue to shorten. The precise timing of this reversal appears to be flexible and has been advancing in recent decades, potentially due to faster development driven by climate warming (
Zohner
et al
., 2023
). For temperate trees, the key outcomes of development are production of viable seeds, cessation of primary and secondary growth, and maturation of perennating tissues including leaf or flower buds (completion of bud set) before the onset of frost (
Rohde & Bhalerao, 2007
;
Tanino
et al
., 2010
;
Cooke
et al
., 2012
). Because opportunities for development will vary among years, the timing of the temperature effect reversal should be flexible. However, this flexibility has yet to be demonstrated in processes directly related to development such as autumn bud set.
In this study, we investigate how air temperature changes around the summer solstice affect end-of-season (EOS) timing in European beech (
Fagus sylvatica
L.), using bud set and leaf senescence as key physiological markers of autumn phenology (
Singh
et al
., 2017
;
Zohner & Renner, 2019
;
Mariën
et al
., 2021
). We ask three main questions: i) How does cooling before vs. after the solstice affect EOS timing? ii) How do diel (daytime vs. nighttime) temperature changes differ in their effects? iii) Does the timing of leaf-out modulate the reversal of temperature effects on autumn phenology? Diel temperature variations are relevant because trees typically grow more at night when temperatures and water deficit are lower (
Steppe
et al
., 2015
;
Mencuccini
et al
., 2017
;
Zweifel
et al
., 2021
). Thus, developmental effects mediated by temperature may be especially pronounced during nights. Moreover, long-term trends in climate show that nighttime temperatures rose faster than daytime temperatures from 1950 to 2004 (Vose et al., 2005), though this trend may have shifted since 1991 (Zhong et al., 2023). To address the third question, we experimentally manipulated spring conditions to create early-leafing and late-leafing individuals within the same growing season, allowing us to assess whether slowed early-season development postpones the reversal date of temperature effects on autumn phenology (
Fig. 1
). As depicted in our conceptual model (
Fig. 1
), trees are dynamic systems that continuously integrate and adapt their physiological responses to environmental conditions over time. We therefore expected their temperature responses to differ both across months (June-August) and between day and night.
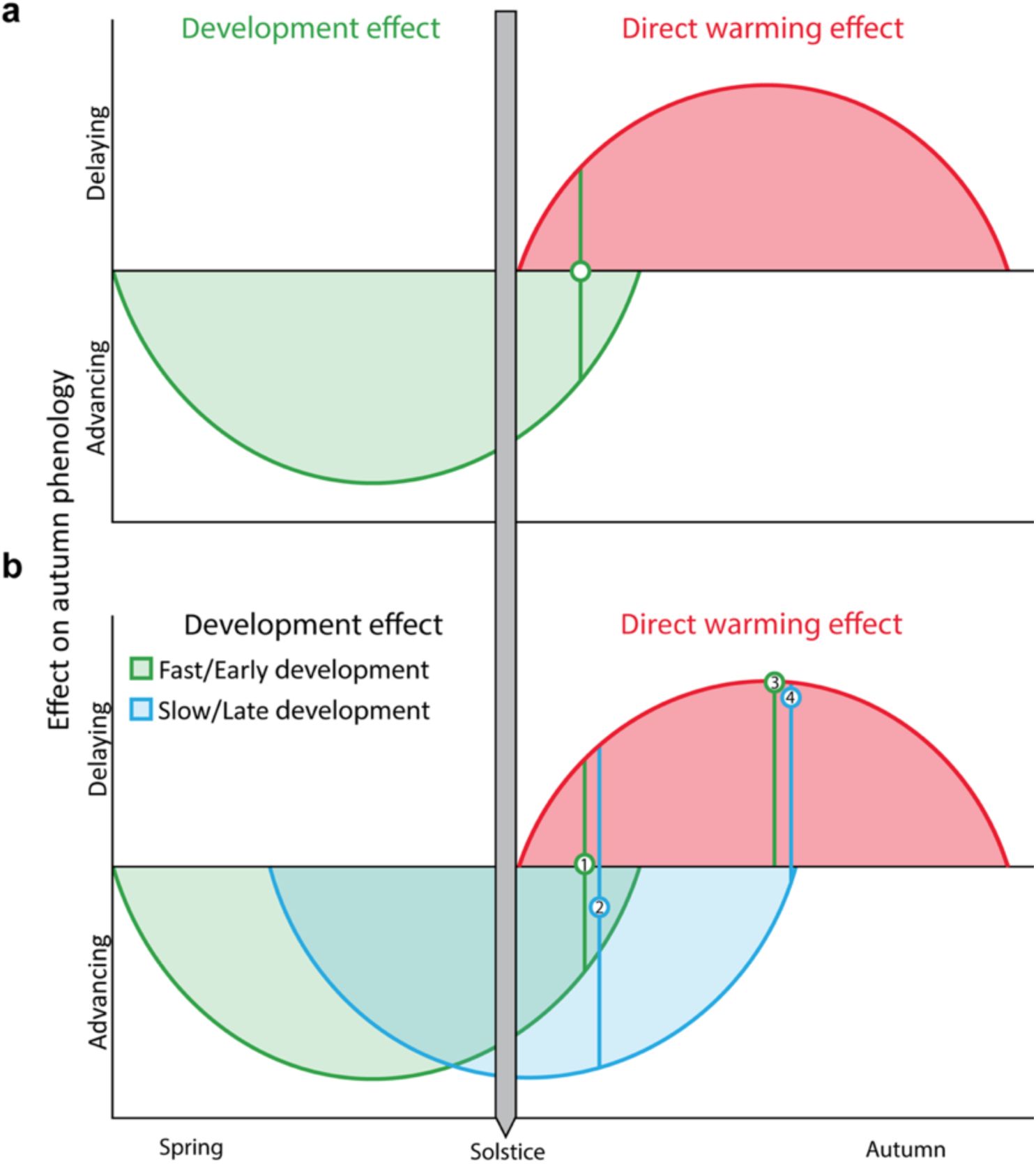
Conceptual model of autumn phenological responses of temperate trees to early-season development and late-season temperature.
Autumn phenology, represented in this study by the timing of primary growth cessation (bud set) and leaf senescence (50% loss of leaf chlorophyll content), is influenced by two opposing factors: early-season development and late-season temperature.
a)
In our model, early-season development, which is driven by temperature, has an advancing effect on autumn phenology that lasts until shortly after the summer solstice (green curve). Higher temperatures cause trees to complete their annual life cycles faster, allowing them to set buds and senesce leaves. After the summer solstice, as days shorten, trees become increasingly sensitive to cooling conditions, so late-season warming slows the progression of bud set and senescence, delaying autumn phenology (red curve). The compensatory point, where the advancing effect of early-season development is balanced by the delaying effect of late-season warming, is represented by the green circle.
b)
According to the model, the timing of this effect reversal is flexible and varies between years based on the speed of development. When development is slow or starts late (blue curve), the effect reversal occurs later than under fast or early development (green curve). Therefore, shortly after the solstice, the effect of temperature on autumn phenology differs between fast/early and slow/late developing individuals. For example, point 1 (green circle) shows no net temperature effect on phenology in fast/early trees. By contrast, point 2 (blue circle) shows that in slow/late developing trees, warmer temperatures shortly after the solstice (in July) still advance autumn phenology. However, as the growing season progresses and days shorten, trees become more responsive to cooling regardless of prior developmental speed. By August, both fast/early and slow/late trees should therefore exhibit similar phenological responses, with warming consistently delaying autumn phenology (points 3 and 4).
Materials and Methods
Selected species
Fagus sylvatica
L. has a large temperate European distribution, is highly economically and ecologically important, and is likely threatened by climate change (
Gessler
et al
., 2006
). Therefore, improving our understanding of how
F. sylvatica
’s growing season is modulated is vital to the management of European forests. Moreover,
F. sylvatica
has frequently been used as a key species to study temperate tree phenology (
Dittmar & Elling, 2006
;
Vitasse
et al
., 2011
;
Zohner & Renner, 2019
;
Zani
et al
., 2020
;
Zohner
et al
., 2023
;
Mariën
et al
., 2025
;
Švik
et al
., 2025
).
Experiment 1
To test the antagonistic effects of early-season development and late-season air temperature on autumn phenology, we set up an experimental population (n = 267) of 40-60 cm tall European beech (
F. sylvatica
) trees in Zurich, Switzerland in 2023. The trees were sourced from a local nursery, and each tree was placed individually in a 20 L plastic pot containing a 1:1:1 sand / peat / organic soil mixture with a Nitrogen (N) concentration of ∼65 g m
−3
, a Phosphate (P
2
O
5
) concentration of ∼140 g m
−3
, and a Potassium (K
2
O) concentration of ∼400 g m
−3
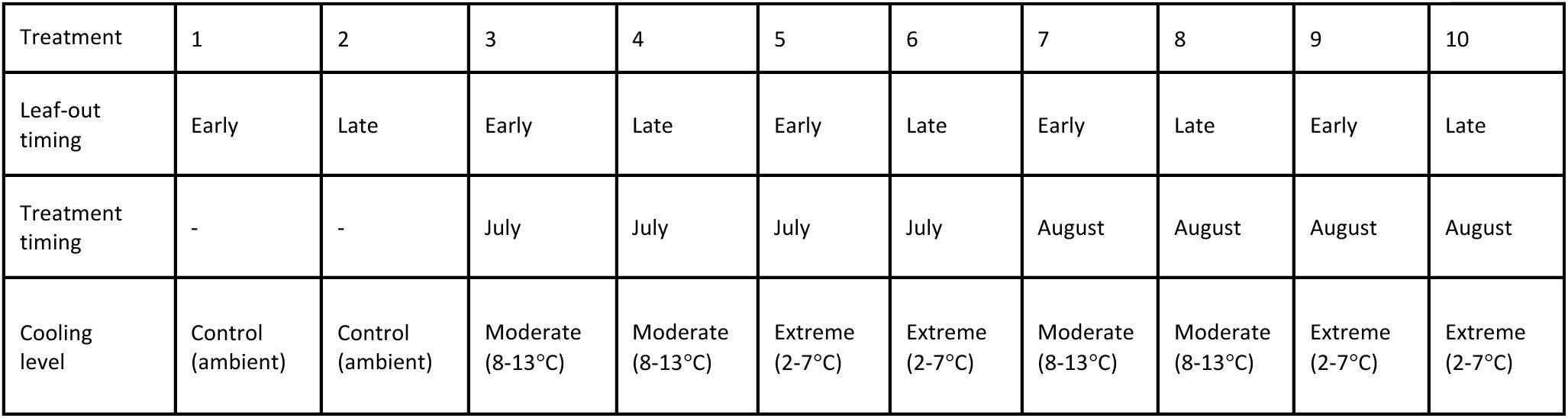
. Individual trees were assigned randomly to one of 10 treatment groups (26 ≤ n ≤ 27). Treatment groups were cooled at different times of the year, using different cooling levels (
Fig. 2
,
Table 1
). To arrest spring development and thereby generate early-leafing and late-leafing individuals, half of the experimental population was placed outside under ambient conditions, while the other half was cooled in climate chambers from 4 April to 24 May. The chambers were set to a low of 2°C at night and a high of 7°C during the day, following a simulated day-night cycle of temperature and light availability (13 h photoperiod at ∼4,300 lux). These cold conditions were not intended to mimic natural European conditions, but rather to slow spring development as much as possible without damaging the trees. Temperatures around 5°C are associated with low rates of growth and development processes such as cell division, expansion and maturation (
Körner, 2003
;
Tumajer
et al
., 2021
). From 24 May to 21 June, all trees were kept outside under ambient conditions in a randomised block design. All trees were monitored to observe their individual leaf-out dates, which was defined as the date when > 50% of their leaves had unfolded, corresponding to BBCH15 (
Capdevielle-Vargas
et al
., 2015
).
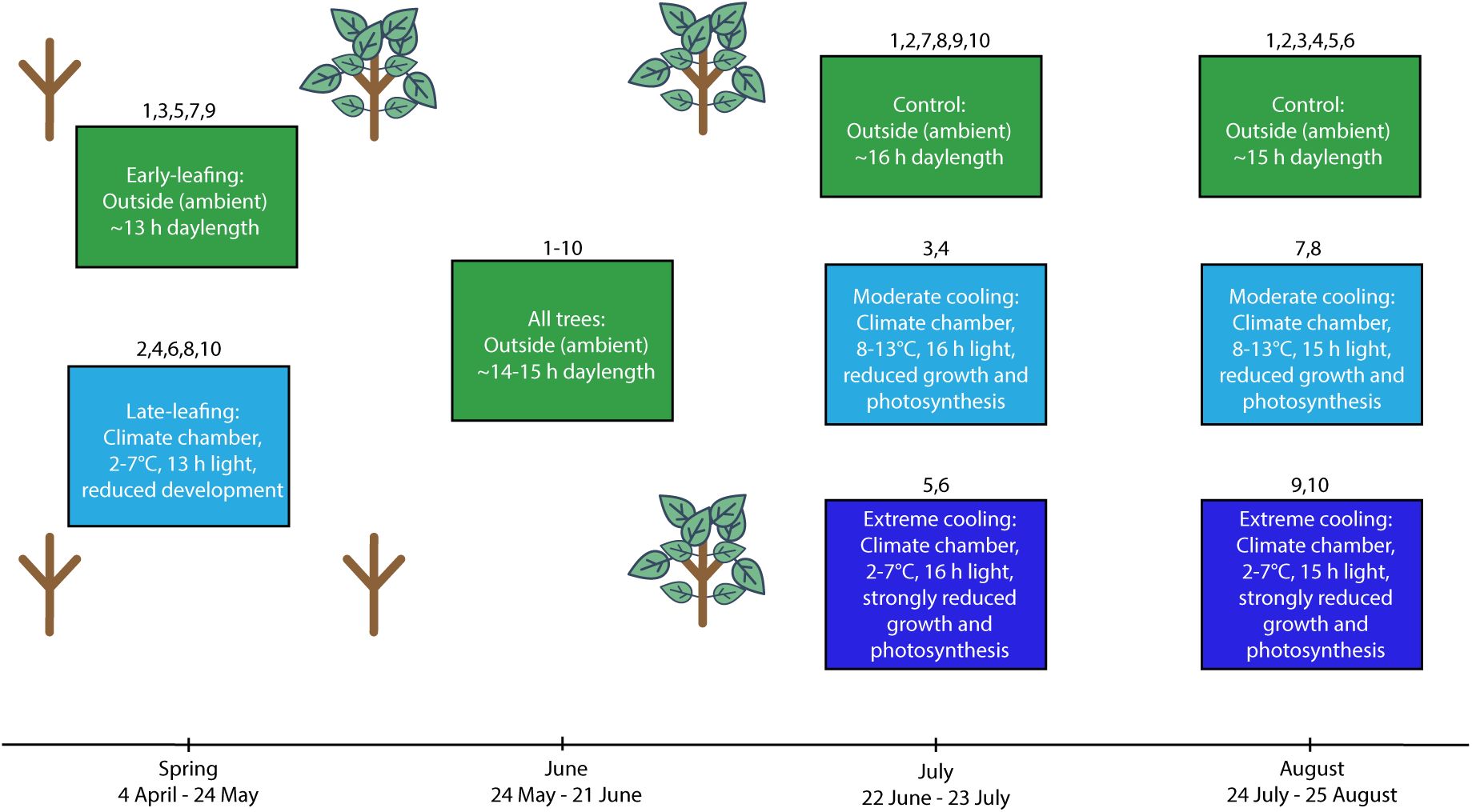
Depiction of the experimental timeline and settings for experiment 1.
Each box corresponds to a specific treatment block at that point in time. The numbers inserted above the boxes refers to the treatment group (see
Table 1
for details). Each box contains information on the location of the trees, the specific conditions they were under and the intended physiological effects of those conditions. The sapling graphics highlight differences in early-season developmental progression for the early-leafing and late-leafing groups. Following the August treatments all trees were placed outside under ambient conditions in a randomised block design.
Description of the temperature treatments applied in experiment 1.
July treatments were from 22 June to 23 July. August treatments were from 24 July to 25 August. Ambient means outside under natural conditions. Temperature ranges indicate the daily minimum and maximum temperatures experienced by trees inside climate chambers. The early-leafing and late-leafing trees were experimentally generated by placing potted trees in climate chambers from 4 April to 24 May and cooling them to 2°C at night and 7°C during the day to arrest spring development.
During the summer treatments, control trees were placed outside under natural ambient conditions. We did not use climate chambers for the control groups because warm conditions in these chambers can introduce unique physiological stressors—such as aphid infestations— not present in cold chambers (
Bezemer
et al
., 1998
). In experiment 2, however, we included an additional chamber control to test for potential chamber effects. Although we observed higher aphid abundances in these warm control chambers, chamber exposure itself had no detectable effect on autumn phenology. This strengthens the inference that the phenological shifts observed in our treatments reflect the intended temperature manipulations. Nevertheless, for experiment 1, where chamber controls were not included, we cannot entirely rule out the possibility that chamber conditions exerted some unintended influence on treated trees compared with ambient controls.
The July treatments took place directly after the summer solstice from 22 June to 23 July. Treated trees were placed in climate chambers set to a low of 2°C at night and a high of 7°C during the day or a low of 8°C at night and a high of 13°C during the day depending on their cooling level (extreme and moderate, respectively). Trees in the chambers experienced a photoperiod of 16 h at ∼7,300 lux. The extreme cooling was designed to severely impair cell division, expansion, and maturation as described above. Additionally, under this temperature regime, photosynthesis should be reduced by > 40% (
Körner, 2006
). The moderate cooling regime, which was still much colder than the ambient conditions, should have also impaired growth and development as well as reduced photosynthesis by at least 30% (
Körner, 2003
,
2006
;
Lenz
et al
., 2014
). These settings aimed to generate large temperature differences between treatments and maximise our ability to detect the mechanism underlying the solstice switch, rather than to represent naturally occurring conditions. The August treatments took place from 24 July to 25 August under the same conditions as the July treatments except with a 15 h photoperiod. For the remainder of the experiment, all trees were kept outside in a randomised block design. Throughout the experiment, all trees were watered frequently to ensure constant water supply (
Fig. S1
).
To observe the effects of our treatments on the development of overwintering buds, we monitored bud growth to derive bud set dates, our marker for the cessation of primary aboveground development and the beginning of autumn phenology, for all trees. On each tree, the terminal bud of the primary shoot and the terminal bud on a random lateral stem were selected and tagged for measurement. Each selected bud was measured to 0.01 mm precision using a digital calliper (CD-P8”M, Mitutoyo Corp, Japan). We measured all buds on a regular basis from 4 July to 2 November. Bud set was defined as the date when each bud reached 90% of its own maximum length, which is considered to be an indicative stage of aboveground primary growth cessation (
Signarbieux
et al
., 2017
;
Zohner & Renner, 2019
). Bud lengths were linearly interpolated between measurement dates to derive the date at which they reached the 90% threshold.
To obtain a more holistic perspective of autumn phenology, we also derived leaf senescence dates from leaf spectral index measurements taken using a SPAD-502 Plus (Soil Plant Analysis Development, Minolta Camera Co., Ltd, Tokyo, Japan). We measured nine leaves per individual (three each from the top, middle and bottom of the crown) on a monthly basis in summer, every other week in September, and on a weekly basis from October to mid-December. We removed measurements taken on 7 December from the analysis as they were unreasonably high, likely due to false readings caused by extensive leaf browning. SPAD readings were then converted to total chlorophyll content (
Chl
in µg/g fresh weight) using an empirically established equation for
Fagus sylvatica
leaves (
Percival
et al
., 2008
):
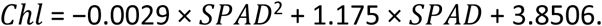
The chlorophyll content between two consecutive measurement dates was estimated using linear interpolation. For each treatment and measurement date, we removed individual chlorophyll measurements that were more than 1.5 × the interquartile range below the lower quartile or above the upper quartile. As an additional cleaning step, we completely removed any trees that had more than one data point removed in the last step. Finally, we calculated individual leaf senescence dates as the day-of-year when chlorophyll content last fell below 50% of the observed peak chlorophyll content.
Experiment 2
To observe the effects of pre- and post-solstice daytime and nighttime temperature on autumn phenology, we set up an experimental population (n = 180) of four-year-old
Fagus sylvatica
trees in Zurich, Switzerland in 2022. The trees were sourced from a local nursery and each tree was placed individually in a 20 L plastic pot containing a 1:1:1 sand / peat / organic soil mixture with a Nitrogen (N) concentration of ∼65 g m
−3
, a Phosphate (P
2
O
5
) concentration of ∼140 g m
−3
, and a Potassium (K
2
O) concentration of ∼400 g m
−3
.
The ambient control treatment consisted of 36 trees exposed to natural ambient conditions. The remaining eight treatments were each applied to 18 trees and included cooling in climate chambers with simulated ambient day length (16 h) and light intensity (∼6,900 lux). The pre-solstice cooling treatments were applied between 22 May and 21 June. The post-solstice cooling treatments were applied between 22 June and 21 July. The pre- and post-solstice treatments included four levels each: Chamber control, where the trees were continuously subjected to 20°C; Day cooling, where trees were subjected to 8°C in the daytime and 20°C at night; Night cooling, where trees were subjected to 20°C in the day and 8°C at night; Full-day cooling, where trees were continuously subjected to 8°C (
Fig. 3
,
Table 2
). Due to warm temperatures, chamber control trees were subject to an increase in aphid abundance, however this did not alter their bud set timing (see data analyses). After treatment, all trees were placed in a randomised block design outside under ambient conditions. Soil moisture content was regulated by frequent watering. We measured all buds on a weekly basis from 25 August to 3 November following the same methodology as in experiment 1. We also measured leaf-level CO
2
assimilation rates during the pre-solstice treatment window (see
Zohner
et al
., 2023
for methodology), and derived leaf senescence dates following experiment 1.
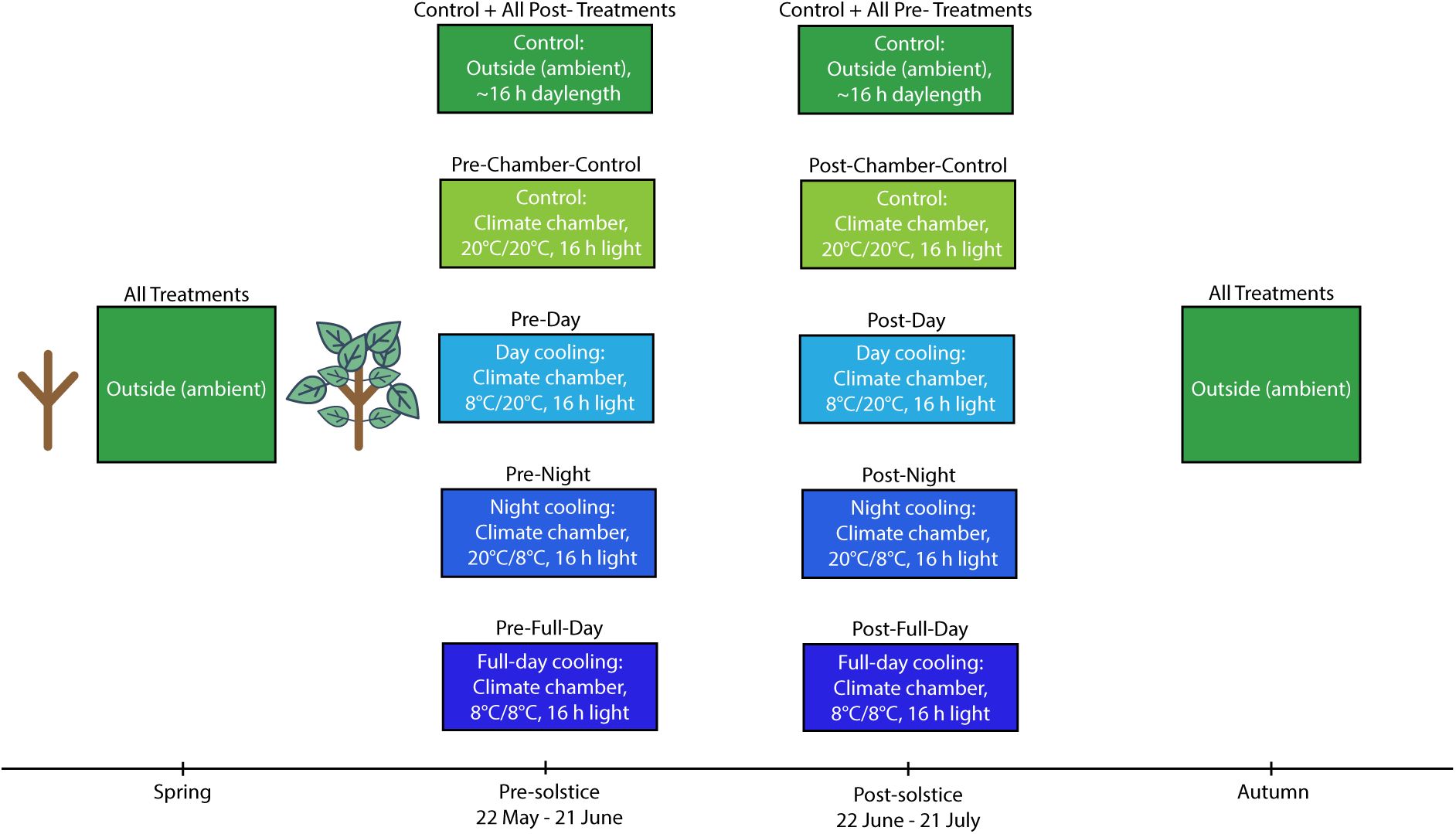
Depiction of the experimental timeline and settings for experiment 2.
Each box corresponds to a specific treatment block at that point in time. The text inserted above the boxes refers to the treatment group (see
Table 2
for details). Each box contains information on the location of the trees and the specific conditions they were under. The sapling graphics indicate that all trees had equal opportunities for early-season development.
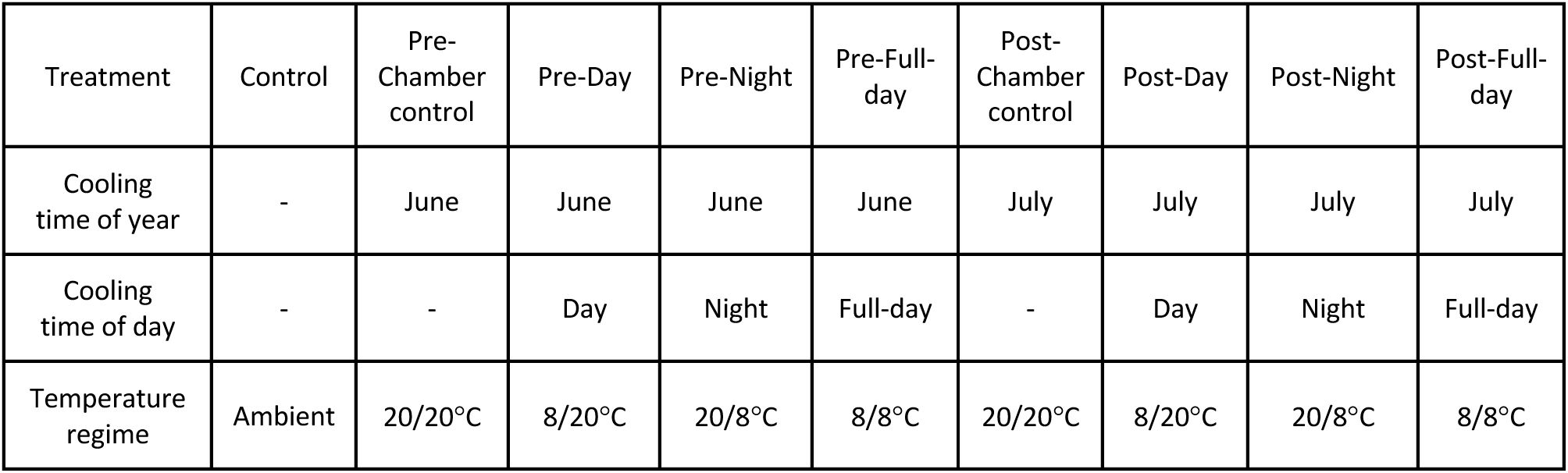
Description of the temperature treatments applied in experiment 2.
June treatments were from 22 May to 21 June. July treatments were from 22 June to 21 July. Ambient means outside under natural conditions. The remaining temperature regimes are in the format day/night and refer to the temperatures applied to trees in climate chambers
Data analyses
For experiment 2, we performed one-way ANOVAs that showed no significant differences between the end-of-season dates of the ambient, pre-chamber and post-chamber control groups (for bud set:
F
2,136
= 0.346, p = 0.708, see
Fig. S7
; for leaf senescence:
F
2,63
= 0.931, p = 0.399). Therefore, for the following analyses we treated all three control treatments as one control treatment with 72 trees. For both experiments we ran linear models using treatment and bud type (apical vs. lateral) as predictors and bud set day-of-year, absolute bud growth or relative bud growth as the response variable. When modelling leaf senescence day-of-year, we used treatment as the sole predictor. For experiment 1, effect sizes were calculated in comparison to the corresponding control treatment, i.e., all early-leafing treatments were compared to the early-leafing control treatment, and all late-leafing treatments were compared to the late-leafing control. Absolute and relative bud growth were calculated as:
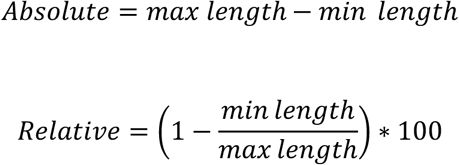
Where:
min length
= the first measured bud length,
max length
= the measured bud length when the bud first surpassed 90% of its final length. To determine the sensitivity of autumn bud set timing to spring leaf-out timing, we ran a linear mixed-effects model using leaf-out day-of-year and bud type as the fixed predictor variables, summer temperature treatment as random effect, and bud set day-of-year as the response variable. We ran the same analysis for leaf senescence timing without the bud type effect.
To quantify the relative contributions of early-season development and late-season temperature to variation in bud set timing in experiment 1, we ran a variance partitioning analysis. We fit a linear model with leaf-out day-of-year, summer cooling treatment (combination of cooling timing and level, e.g. July_Moderate), and bud type as explanatory variables. Variance partitioning was conducted using the
varpart
function in the
vegan
R package (
Oksanen
et al
., 2025
), which decomposes explained variance into unique and shared components. All statistical analyses were performed in R version 4.5.1 (
R Core Team, 2025
).
Results
Experiment 1
Linear modelling showed that across all treatments, late-leafing trees cooled during spring delayed their bud set by 4.6 ± 1.3 days (mean ± 2 SE, p < 0.01). Across all treatment pairings, the mean bud set date for the late-leafing group always occurred later than for the early-leafing group (
Fig. S2
). A linear mixed effects model including leaf-out day-of-year and bud-type as fixed effects and treatment as a random effect revealed that, on average, each day delay in spring leaf-out was associated with a delay of 0.24 ± 0.06 days in bud set timing. Across all treatments, bud type had a small effect, lateral buds set on average 1.1 ± 1.2-days earlier than apical buds (p = 0.06). When modelling only the early-leafing groups, bud type had no discernible effect (0.04 ± 1.64 days earlier for lateral buds, p = 0.96). However, when modelling only the late-leafing groups, lateral buds set earlier than apical buds (2.17 ± 1.64 days advancement, p = 0.01).
The effect of July cooling differed strongly between the early- and late-leafing trees: Within the early-leafing group, moderate cooling in July lead to a small, non-significant delay in bud set compared to the ambient July treatment (1.4 ± 2.6 days, p = 0.29). July cooling had a much greater impact on late-leafing trees, leading to a 4.8 ± 2.6-day delay in bud set compared to the ambient July treatment in the late-leafing group (p < 0.01).
In contrast to July cooling, the effects of August cooling did not differ between early- and late-leafing trees, advancing bud set in all treatment groups (
Fig. 4b
). Cooling in August lead to a 4.5 ± 2.6-day (p < 0.01) and a 4.4 ± 2.6-day (p < 0.01) advancement in bud set for early- and late-leafing trees, respectively. The extreme cooling treatments showed similar patterns, although slightly less clearly (
Fig. S3
). Variance partitioning revealed that the early-season development effect (leaf-out day-of-year) explained 8.8% of the total variance in bud set timing, the late-season temperature effect (summer cooling treatment) explained 14.6%, and bud type explained 0.3%. Shared variance values were ≍ 0%, indicating no meaningful overlap. Effects of absolute and relative growth on bud set date can be found in the supporting information (Figs. S4-S5). Bud set timing had no effect on final bud length (
Fig. S6
).
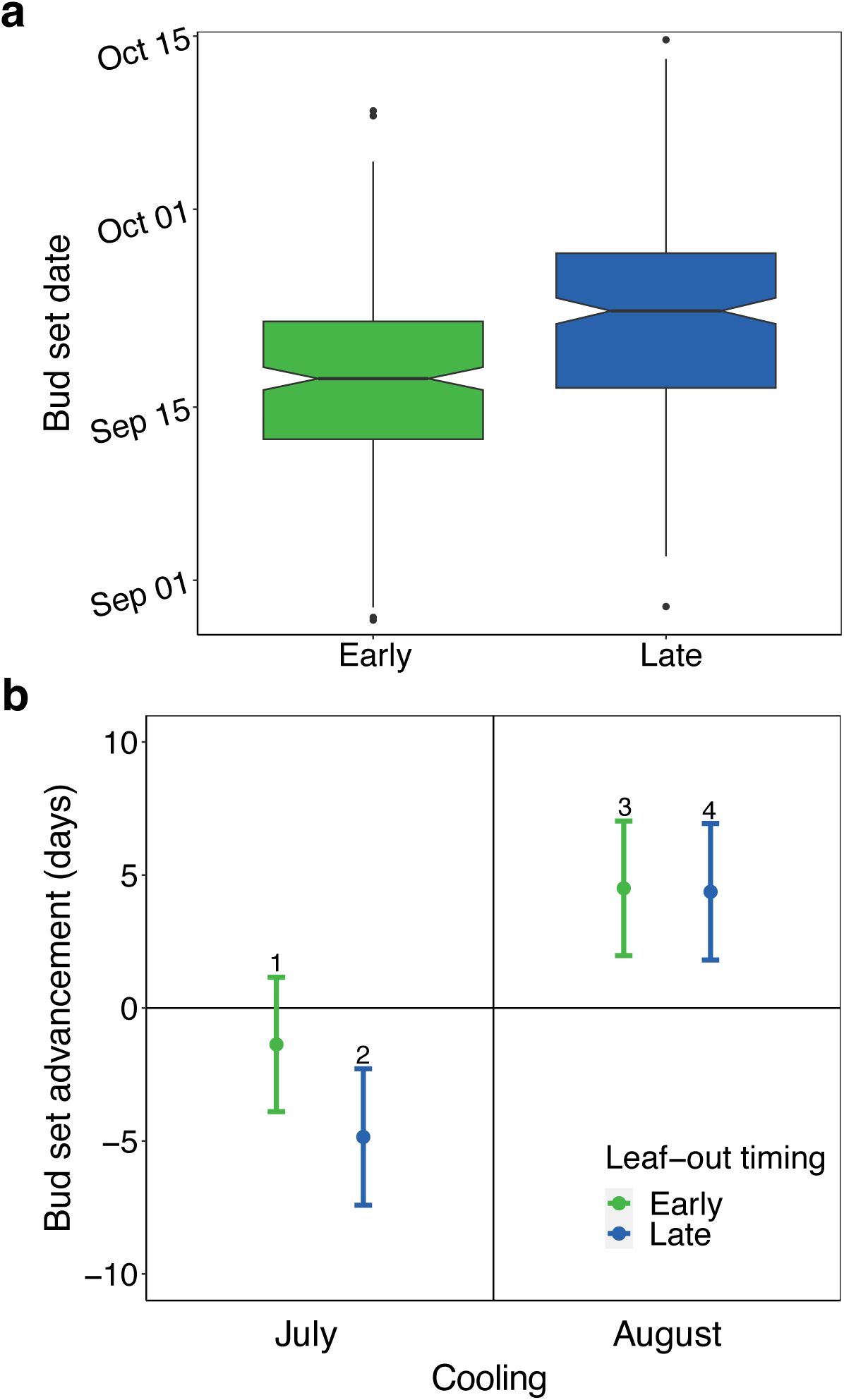
Effects of early-season development and late-season temperature on the timing of autumn bud set in
Fagus sylvatica
(experiment 1).
a)
Bud set dates for early- (green) and late- (blue) leafing trees including all treatments. Late-leafing trees were cooled (2-7°C) in climate chambers from 4 April to 24 May to arrest their development and delay leaf-out.
b)
Effects of July (22 June to 23 July) and August (24 July to 25 August) moderate cooling (8-13°C) on bud set date for early- (green) and late- (blue) leafing trees (see
Fig. S3
for extreme cooling effects). Analyses show effect size means ± 95% confidence intervals from linear models, including treatment and bud-type (apical vs lateral) as predictors. Early-leafing effects are calculated against the early-leafing control and late-leafing effects are calculated against the late-leafing control. The bud type effect is not shown. Number labels (1-4) above each point are shown to aid comparison between points 1-4 in the conceptual model (
Fig. 1b
) and the observed effects. Positive values indicate advances in bud set and negative values indicate delays.
Experiment 2
Linear modelling showed that pre-solstice full-day (day and night) cooling delayed autumn bud set by 4.1 ± 3.6 days (mean ± 2SE, p = 0.02) (
Fig. 5
). Pre-solstice nighttime cooling had a similar effect, delaying bud set by 4.2 ± 3.6 days (p = 0.02). By contrast, pre-solstice daytime cooling had no significant effect on bud set (0.1 ± 3.7 days, p = 0.94). Post-solstice full-day cooling advanced autumn bud set by 5.2 ± 3.6 days (p < 0.01). Similarly, post-solstice daytime cooling advanced bud set by 5.3 ± 4.3 days (p = 0.01). Conversely, post-solstice nighttime cooling delayed bud set by 3.8 ± 3.5 days (p = 0.03). Across all treatments, lateral buds set considerably earlier than apical buds (9.4 ± 2.08 days, p < 0.01). Effects of relative growth on bud set date can be found in the supporting information (Figs. S8).
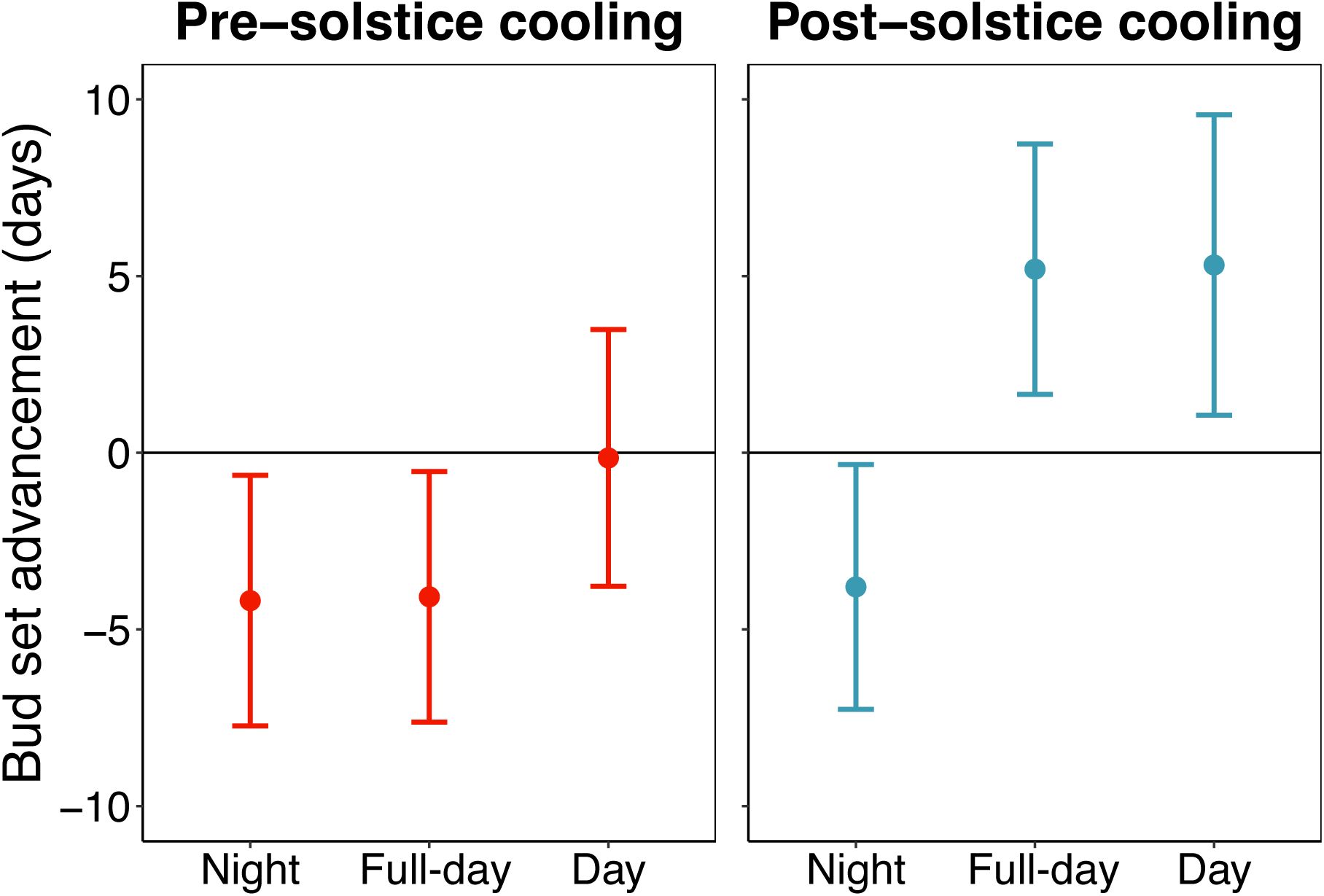
The pre- and post-solstice effects of night, full-day and day cooling on the timing of autumn primary growth cessation in
Fagus sylvatica
(experiment 2).
Effects of pre-solstice (22 May to 21 June) and post-solstice (22 June to 21 July) cooling on bud set date. Full-day cooling trees were continuously cooled to 8°C, day cooling trees were cooled to 8°C in the day and kept at 20°C at night, night cooling trees were cooled to 8°C at night and kept at 20°C in the day. Analyses show effect size means ± 95% confidence intervals from linear models, including treatment and bud-type (apical vs lateral) as predictors. The bud type effect is not shown.
Comparison of phenological metrics
Analyses using leaf senescence (50% drop in chlorophyll content) as the phenological marker produced results that were overall consistent with the bud set analyses. For example, in experiment 1, senescence was delayed by 4.22 ± 1.61 days in late-leafing trees and each day delay in spring leaf-out led to a 0.22 ± 0.08-day delay in leaf senescence, closely matching with bud set. Across both experiments the observed patterns were largely the same, although the strengths of each effect differed slightly, and overall, the effects on leaf senescence tended to be weaker (Figs. S9-10). Out of all treatments, opposing responses from bud set and leaf senescence timing were only observed in the early-leafing extreme August cooling group (Treatment 9, bud effect = 1.9 ± 2.59 days advancement, p = 0.14, leaf effect = 1.77 ± 3.53 days delay, p = 0.32) from experiment 1, and the post-full-day cooling group from experiment 2 (bud effect = 5.2 ± 1.81 days advancement, p < 0.01, leaf effect = 1.66 ± 3.34 days delay, p = 0.32). Out of 66 pairwise comparisons of estimated marginal means (every treatment compared against every other treatment in the same experiment), 55 (83%) showed directional agreement between bud set and leaf senescence effects (
Fig. S11
, adjusted
R
2
= 0.49).
Discussion
In this study, we conducted experiments on European beech to test how bud set—a marker of primary growth cessation—and leaf senescence respond to monthly and diel temperature changes around the summer solstice. We found large differences in the responses between trees subjected to daytime versus nighttime cooling before and after the solstice (
Fig. 5
) and between early- and late-leafing trees (
Fig. 4
). These differences suggest that early-season developmental timing not only influences the timing of autumn phenology
per se
but also plays a critical role in determining the timing of the summer solstice effect reversal. Therefore, our findings support our conceptual model (
Fig. 1
), highlighting development as a key factor underpinning the Solstice-as-Phenology-Switch hypothesis. In the following sections, we discuss the effects of pre-solstice and post-solstice air temperature on development, along with the distinct impacts of daytime versus nighttime temperature variations.
Early-season development rates alter responses to late-season temperature
Our trans-solstice climate manipulation experiments showed that trees delayed the end of their growing season in response to slow/late early-season development (delayed spring leaf-out). Regardless of the summer cooling treatments, late-leafing trees consistently set buds and senesced leaves after their early-leafing counterparts (
Fig. S2
), with each day delay in spring leaf-out delaying bud set by an average of 0.24 days and senescence by 0.22 days. This is in line with previous studies demonstrating a tight linkage between within-year variations in spring and autumn phenology (
Fu
et al
., 2014
;
Keenan & Richardson, 2015
;
Signarbieux
et al
., 2017
), likely governed by developmental constraints (
Zohner
et al
., 2023
), buildup of water and nutrient stress (
Paul & Foyer, 2001
;
Buermann
et al
., 2018
;
Bigler & Vitasse, 2021
), or leaf aging (
Lim
et al
., 2007
).
Despite the importance of early-season development, summer cooling treatment uniquely explained more variance in bud set timing than leaf-out day-of-year (Adjusted
R
2
= 0.15 and 0.09, respectively). July cooling induced a delay in bud set dates 3.4 times greater in late-leafing trees compared to early-leafing ones (4.8 versus 1.4 days delay), which agrees with the expectations derived from our conceptual model (
Fig. 1
). As we had expected, the natural interannual variation in early-season development causes year-to-year differences in the point at which cool temperatures switch from delaying autumn phenology to advancing it. This offers a physiological explanation for the advancement in the effect reversal timing observed between 1966 and 2015 in
Fagus sylvatica
,
Aesculus hippocastanum
,
Quercus robur
, and
Betula pendula
(
Zohner
et al
., 2023
; because control trees did not experience chamber time, we cannot entirely rule out some influence of this factor on our comparisons).
August cooling induced comparable advances in bud set timing in both early- and late-leafing trees (4.4-4.5 days). The diminishing difference in cooling responses between the two groups from July to August suggests that the influence of early-season development weakens as individuals approach completion of their developmental requirements. By August, phenology appears primarily governed by the delaying effect of warm air temperature (see direct warming effect in
Fig. 1b
).
Two mechanisms could underlie this shift: shortening daylengths and declining photosynthetic assimilation, the latter of which has been linked to accelerated senescence (
Krieger-Liszkay
et al
., 2019
). However, an experiment that reduced August photosynthetic rates in beech by 52-72% – either through cooling or shading – showed that photosynthetic assimilation alone cannot account for autumn phenology (
Zohner
et al
., 2023
). Despite similar reductions in photosynthesis, August cooling advanced senescence, while August shading delayed it, revealing contrasting outcomes. This suggests that photosynthetic assimilation cannot have been the driver of the observed phenological responses in beech. Instead, the observed change in temperature responses is more likely regulated by daylength (
Kramer, 1936
;
Petterle
et al
., 2013
;
Singh
et al
., 2021
). As days shorten, trees become increasingly responsive to cooling (
Delpierre
et al
., 2009
;
Körner
et al
., 2016
). Together with the waning influence of developmental carry-over, this can explain why by August trees responded similarly to cooling regardless of their earlier developmental trajectories (
Fig. 1
). These findings highlight the critical interaction between photoperiod and temperature in shaping the autumn phenology of
Fagus sylvatica
.
Effects of daytime vs. nighttime temperature
Our experiments showed that daytime and nighttime temperatures had different effects on autumn phenology before and after the summer solstice: Before the solstice, daytime cooling had no impact on the timing of bud set, while nighttime cooling and full-day cooling delayed bud set by 4.2 and 4.1 days, respectively (
Fig. 5
). This pattern was similar when using leaf senescence as the end-of-season marker. Because experimental cooling reduced photosynthesis by 69-83% across all treatments (
Fig. S12
), these effects are unlikely to be explained by changes in carbon assimilation. Instead, they highlight the importance of developmental processes—such as cell division and expansion—which mostly occur at night (
Steppe
et al
., 2015
;
Mencuccini
et al
., 2017
;
Zweifel et al., 2021
;
Fig. 6a
). The developmental arrest induced by night cooling—and therefore also by full-day cooling—likely slowed growth and maturation, effectively extending the growing season.
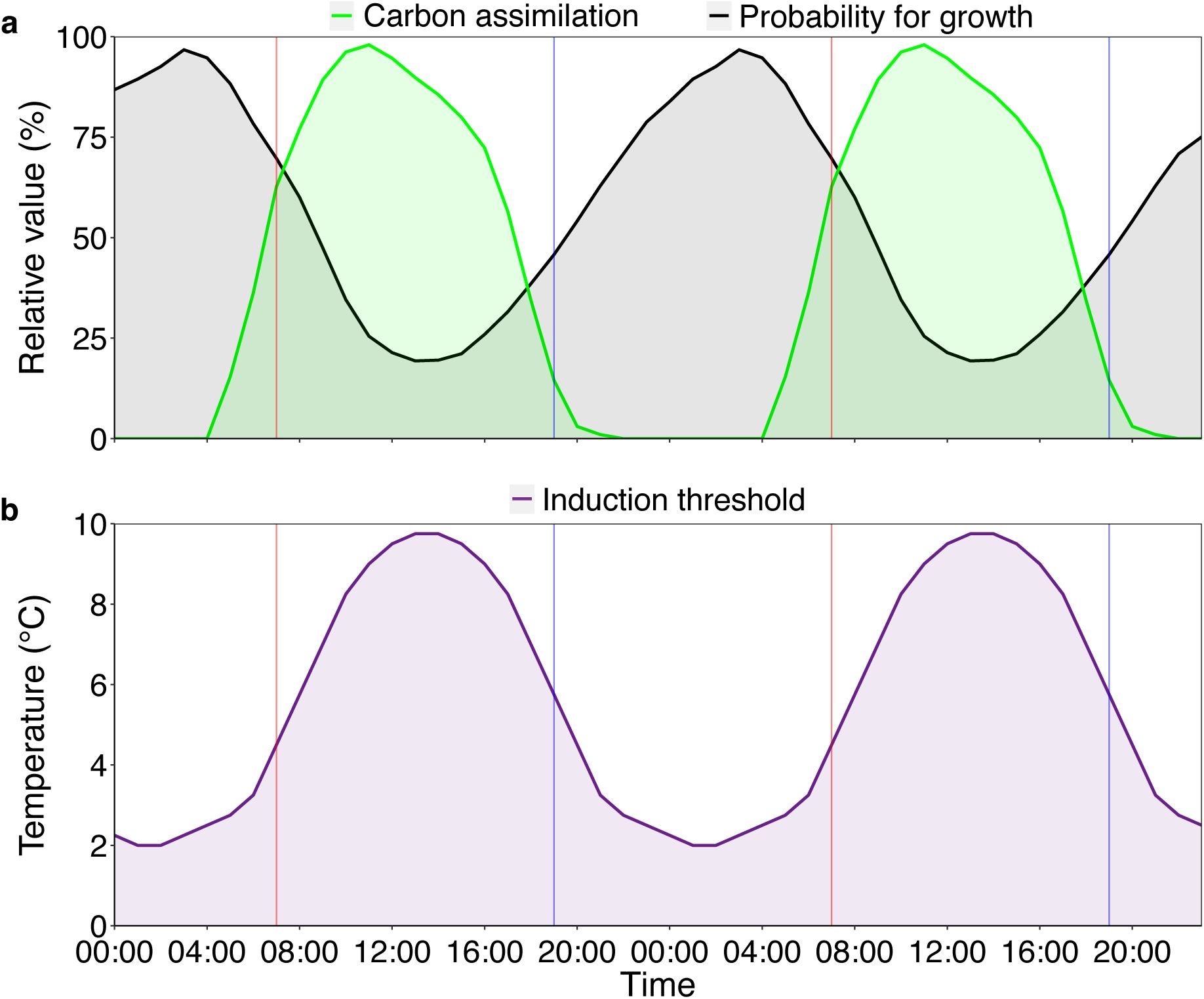
Diel patterns of relative growth, photosynthetic rate and theoretical thresholds for cold-induced bud set in
Fagus sylvatica
.
Vertical red lines indicate the start of the day, and vertical blue lines indicate the start of the night, marking the boundaries between the 12-hour treatment windows used in experiment 2.
a)
The green curve shows relative carbon assimilation rate, the raw values were taken from the literature in a study that measured assimilation rates under controlled conditions (
Urban
et al
., 2014
). Values were linearly interpolated between measurements then converted to a percentage of the peak value. Finally, the curve was smoothed by taking the running mean of the target value, the previous value and the following value. The black curve shows the relative probability for growth, the raw values were taken from the literature in a study that measured growth rates in the field (
Zweifel
et al
., 2021
), then processed in the same way as the assimilation data. At night, low temperatures slow the trees’ developmental processes, possibly leading to delayed tissue maturation, while low temperatures during the day reduce photosynthetic activity.
b)
The Diel Cooling Hypothesis: As days shorten after the solstice, autumn bud set becomes increasingly responsive to cooling. Temperatures below a certain threshold induce overwintering responses, advancing autumn phenology. Our results indicate that daytime cooling of 8°C is below this threshold. However, because daily temperatures reach their minimum during the night, trees’ induction thresholds should be lower at night than during the day. Post-solstice night-time cooling of 8°C may thus have delayed bud set by slowing development rather than inducing overwintering responses.
After the solstice, daytime and nighttime cooling of 8°C elicited opposite responses in both phenological markers, with weaker effects on senescence. Trees subjected to post-solstice daytime cooling and full-day cooling set their buds earliest, on average more than five days earlier than controls. As days shorten, the temperature threshold inducing this growth cessation must be lower at night, otherwise trees would risk prematurely ending their growing season due to the daily minimum temperatures occurring at night (
Fig. 6b
). This may explain why post-solstice nighttime cooling delayed, rather than advanced, autumn phenology: Cooling of 8°C was sufficient to trigger overwintering responses—growth cessation and maturation of perennating tissues—when applied during the day but insufficient when applied at night (see
Fig. S13
for a conceptual schematic).
An alternative explanation is that daytime cooling indirectly triggered dormancy induction by suppressing photosynthesis (
Fig 6a
), under the assumption that trees prioritise dormancy once photosynthesis is no longer possible (
Körner
et al
., 2016
). Though, as discussed above, late-season photosynthesis appears to assert little control over the autumn phenology of
F. sylvatica
. Taken together, these results support a Diel Cooling Hypothesis, in which the impact of cooling depends on whether it occurs during the day or at night, because the temperature thresholds that trigger growth cessation differ between daytime and nighttime conditions.
Nighttime cooling always led to a delay in bud set of approximately four days (
Fig. 5
). As trees mainly grow at night (
Steppe
et al
., 2015
;
Mencuccini
et al
., 2017
;
Zweifel
et al
., 2021
), colder nighttime temperatures likely slowed down key developmental processes such as meristematic activity, tissue expansion and maturation, which in turn delayed primary growth cessation (
Fig. 6
). Similar responses may occur in other temperate tree species: Cold autumn nights have been shown to delay growth cessation and slow bud development in
Populus
,
Pinus
and
Picea
species (
Kramer, 1956
;
Malcolm & Pymar, 1975
;
Kalcsits
et al
., 2009
). Moreover, the summer solstice effect reversal has been observed consistently across Northern Hemisphere temperate forests (
Zohner
et al
., 2023
). Further experiments should explicitly test these diel responses in other temperate tree species.
Support for the Solstice-as-Phenology Switch Hypothesis
The Solstice-as-Phenology-Switch hypothesis posits that changes in temperature before and after the summer solstice have opposite effects on the autumn phenology of northern forest trees (
Zohner
et al
., 2023
). The hypothesis does not assume that the reversal is fixed precisely on June 21, but rather that it occurs around the solstice, when photoperiod cues shift trees from accelerating to decelerating their seasonal development. Our conceptual model (
Fig. 1
) explicitly incorporates this flexibility by showing how the timing of the reversal depends on developmental speed: individuals that develop more slowly (or leaf out later) cross the compensatory point later in the summer, whereas fast-developing individuals reach it earlier.
Our experiments support this framework: pre-solstice full-day cooling delayed bud set, whereas post-solstice full-day cooling advanced it, with differences between early- and late-developing individuals consistent with the model. Moreover, the contrasting impacts of daytime vs. nighttime cooling demonstrate how diel conditions can further shape when the reversal is expressed. Thus, our findings reinforce the Solstice-as-Switch hypothesis and extend it by showing how flexibility arises from interactions between developmental progression, diel temperature responses, and photoperiod.
In conclusion, earlier primary growth cessation in temperate trees under climate change may result from higher developmental rates under warmer temperatures. In European beech, we show that this effect is predominantly driven by nighttime temperatures, reflecting the fact that growth primarily occurs at night. To better predict future growing season lengths, models must begin to account for the differential impacts of the seasonal and daily temperature cycles that temperate zone trees are physiologically adapted to.
Data availability
The code and data for this study are available at Zenodo (
Rebindaine
et al
., 2025
).
Acknowledgements
C.M.Z. was supported by the SNF Ambizione Fellowship programme (no. PZ00P3_193646) and T.W.C. by DOB Ecology and the Bernina Foundation.
Additional information
Author contributions
D.R. conceived and developed the study, conducted the experiments and analyses, and wrote the manuscript. C.M.Z equally contributed to the conception and development of the study, contributed to the experiments and analyses, and the text. L.M., H.M., Z.W. and Y.Z. contributed to the conception and implementation of the first experiment. R.B. contributed to the conception and implementation of the second experiment. T.W.C. and S.S.R. contributed to the text. All authors provided comments and approved the final manuscript.
Funding
DOB Ecology
Thomas W Crowther
Bernina Foundation
Thomas W Crowther
Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (SNF) (PZ00P3_193646)
Constantin M Zohner
Additional files
References
Photoperiodic regulation of the seasonal pattern of photosynthetic capacity and the implications for carbon cycling
Proceedings of the National Academy of Sciences
109
:8612–8617
PubMed
Google Scholar
Long-term effects of elevated CO2 and temperature on populations of the peach potato aphid Myzus persicae and its parasitoid Aphidius matricariae
Oecologia
116
:128–135
PubMed
Google Scholar
Premature leaf discoloration of European deciduous trees is caused by drought and heat in late spring and cold spells in early fall
Agricultural and Forest Meteorology
307
:108492
Google Scholar
Impacts of climate change on natural forest productivity - evidence since the middle of the 20th century: climate change impacts on forest vegetation
Global Change Biology
12
:862–882
Google Scholar
Widespread seasonal compensation effects of spring warming on northern plant productivity
Nature
562
:110–114
PubMed
Google Scholar
Multiple-year assessment of phenological plasticity within a beech (Fagus sylvatica L.) stand in southern Germany
Agricultural and Forest Meteorology
211–212
:13–22
Google Scholar
The dynamic nature of bud dormancy in trees: environmental control and molecular mechanisms
Plant, Cell & Environment
35
:1707–1728
PubMed
Google Scholar
Modelling interannual and spatial variability of leaf senescence for three deciduous tree species in France
Agricultural and Forest Meteorology
149
:938–948
Google Scholar
Phenological phases of common beech (Fagus sylvatica L.) and their dependence on region and altitude in Southern Germany
European Journal of Forest Research
125
:181–188
Google Scholar
Variation in leaf flushing date influences autumnal senescence and next year’s flushing date in two temperate tree species
Proceedings of the National Academy of Sciences
111
:7355–7360
PubMed
Google Scholar
Larger temperature response of autumn leaf senescence than spring leaf-out phenology
Global Change Biology
24
:2159–2168
PubMed
Google Scholar
Potential risks for European beech (Fagus sylvatica L.) in a changing climate
Trees
21
:1–11
Google Scholar
Beyond source and sink control – toward an integrated approach to understand the carbon balance in plants
New Phytologist
242
:858–869
PubMed
Google Scholar
Changes in autumn senescence in northern hemisphere deciduous trees: a meta-analysis of autumn phenology studies
Annals of Botany
116
:875–888
PubMed
Google Scholar
Magnetic resonance microimaging indicates water diffusion correlates with dormancy induction in cultured hybrid poplar (Populus spp.) buds
Tree Physiology
29
:1269–1277
PubMed
Google Scholar
Net carbon uptake has increased through warming-induced changes in temperate forest phenology
Nature Climate Change
Google Scholar
The timing of autumn senescence is affected by the timing of spring phenology: implications for predictive models
Global Change Biology
21
:2634–2641
PubMed
Google Scholar
Alpine Plant Life
Berlin, Heidelberg
:
Springer
Google Scholar
Significance of Temperature in Plant Life
In:
Morison JIL
Morecroft MD
, editors.
Plant Growth and Climate Change
Wiley
pp. 48–69
Google Scholar
Where, why and how? Explaining the low-temperature range limits of temperate tree species
Journal of Ecology
104
:1076–1088
Google Scholar
Effect of variation in length of day on growth and dormancy of trees
Plant Physiology
11
:127–137
Google Scholar
Some effects of various combinations of day and night temperatures and photoperiod on the height growth of loblolly pine seedlings
Forest Science
3
:45–55
Google Scholar
The impact of photosynthesis on initiation of leaf senescence
Physiologia Plantarum
166
:148–164
PubMed
Google Scholar
Growth and carbon relations of temperate deciduous tree species at their upper elevation range limit
Journal of Ecology
102
:1537–1548
Google Scholar
Leaf Senescence
Annual Review of Plant Biology
58
:115–136
PubMed
Google Scholar
The influence of temperature on the cessation of height growth of Sitka spruce (Picea sitchensis Bong. Carr.)
Silvae Genet
24
:5–6
Google Scholar
Does drought advance the onset of autumn leaf senescence in temperate deciduous forest trees?
Biogeosciences
18
:3309–3330
Google Scholar
Nature’s Master of Ceremony: The Populus Circadian Clock as Orchestrator of Tree Growth and Phenology
npj Biological Timing and Sleep
2
:1–19
PubMed
Google Scholar
An empirical method that separates irreversible stem radial growth from bark water content changes in trees: theory and case studies
Plant, Cell & Environment
40
:290–303
PubMed
Google Scholar
European phenological response to climate change matches the warming pattern: european phenological response to climate change
Global Change Biology
12
:1969–1976
Google Scholar
vegan: Community Ecology Package
CRAN
Sink regulation of photosynthesis
Journal of Experimental Botany
52
:1383–1400
PubMed
Google Scholar
Phenology Feedbacks on Climate Change
Science
324
:887–888
PubMed
Google Scholar
The Potential of a Chlorophyll Content SPAD Meter to Quantify Nutrient Stress in Foliar Tissue of Sycamore (Acer pseudoplatanus), English Oak (Quercus robur), and European Beech (Fagus sylvatica)
Arboriculture & Urban Forestry
34
:89–100
Google Scholar
Daylength mediated control of seasonal growth patterns in perennial trees
Current Opinion in Plant Biology
16
:301–306
PubMed
Google Scholar
Plant phenology and global climate change: Current progresses and challenges
Global Change Biology
25
:1922–1940
PubMed
Google Scholar
R: A Language and Environment for Statistical Computing
Developmental constraints mediate the summer solstice reversal of climate effects on the autumn phenology of European beech
Zenodo
Climate change, phenology, and phenological control of vegetation feedbacks to the climate system
Agricultural and Forest Meteorology
169
:156–173
Google Scholar
Plant dormancy in the perennial context
Trends in Plant Science
12
:217–223
PubMed
Google Scholar
Asymmetric effects of cooler and warmer winters on beech phenology last beyond spring
Global Change Biology
23
:4569–4580
PubMed
Google Scholar
Growing in time: exploring the molecular mechanisms of tree growth
Tree Physiology
41
:657–678
PubMed
Google Scholar
Photoperiod- and temperature-mediated control of phenology in trees – a molecular perspective
New Phytologist
213
:511–524
PubMed
Google Scholar
Diel growth dynamics in tree stems: linking anatomy and ecophysiology
Trends in Plant Science
20
:335–343
PubMed
Google Scholar
Phenological trends of European beech stands along the Carpathian arc: a 20-year MOD13Q1/MYD13Q1 based analysis
European Journal of Remote Sensing
58
:2506576
Google Scholar
Temperature-driven plasticity in growth cessation and dormancy development in deciduous woody plants: a working hypothesis suggesting how molecular and cellular function is affected by temperature during dormancy induction
Plant Molecular Biology
73
:49–65
PubMed
Google Scholar
Phenological sensitivity to climate across taxa and trophic levels
Nature
535
:241–245
PubMed
Google Scholar
Forward Modeling Reveals Multidecadal Trends in Cambial Kinetics and Phenology at Treeline
Frontiers in Plant Science
12
:613643
PubMed
Google Scholar
Impact of elevated CO2 concentration on dynamics of leaf photosynthesis in Fagus sylvatica is modulated by sky conditions
Environmental Pollution
185
:271–280
PubMed
Google Scholar
Assessing the effects of climate change on the phenology of European temperate trees
Agricultural and Forest Meteorology
151
:969–980
Google Scholar
Phenology, ontogeny and the effects of climate change on the timing of species interactions
Ecology Letters
13
:1–10
PubMed
Google Scholar
Increased growing-season productivity drives earlier autumn leaf senescence in temperate trees
Science
PubMed
Google Scholar
Effect of climate warming on the timing of autumn leaf senescence reverses after the summer solstice
Science
381
:eadf5098
PubMed
Google Scholar
Ongoing seasonally uneven climate warming leads to earlier autumn growth cessation in deciduous trees
Oecologia
189
:549–561
PubMed
Google Scholar
Why trees grow at night
New Phytologist
231
:2174–2185
PubMed
Google Scholar
Article and author information
Author information
Version history
Sent for peer review
:
Preprint posted
:
Reviewed Preprint version 1
:
Reviewed Preprint version 2
:
Cite all versions
You can cite all versions using the DOI
10.7554/eLife.107554
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2025,
Rebindaine et al.
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
views
276
downloads
8
citation
1
Views, downloads and citations are aggregated across all versions of this paper published by eLife.





