 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库Functional definition of theairway progenitor field through overlapping compensatory regulators
Abstract
Tubular organs present a common solution to fluid transport in multicellular organisms. They often arise by an initial bulging of flat epithelial progenitor cells, which then undergo branching morphogenesis. Here, we present 3 cooperative programs fully defining the
Drosophila
airway progenitor field and their roles in early morphogenesis linking the radial pattern of the 2-dimensional (2D) field to the proximo-distally patterning of the 3D tubes. We previously showed that extrinsic Hedgehog (Hh) and intrinsic POU-Homeobox TF Ventral-veinless (Vvl)/Drifter/U-turn dominantly drive the transcriptional program toward the distal airway cell identity at the expense of a proximal program specified by the GATA TF
grain
(
grn
). Both programs require the basic-HLH-POU TF
trachealess
(
trh
) (Matsuda et. al, 2015). Whereas
trh
is not essential for primordia invagination, we show that in
hh vvl
double mutants, the oval-shaped primordia frequently remain at the 2D plane, retaining
trh
expression in a
grn
dependent manner. Therefore,
hh
and
vvl
are the principal regulators of progenitor invagination independent of
trh
. Each of the 3 regulators, Trh, Vvl and Grn fulfills only complementary or compensatory functions in transcription and morphogenesis but their combinations functionally define the airway progenitor field. We further provide a comprehensive description for allocating the airway progenitors on the body coordinates, involving dorsal Decapentaplegic/BMP signaling along the dorso-ventral axis and subsequent radial EGFR signaling along the proximo-distal axis. The presence of 3 complementary, regulatory programs in early gene expression and morphogenesis of the simple
Drosophila
airways may reflect the vital needs for respiration, and their influence on the evolution of various strategies in tubular organ development.
Introduction
The reiterative tube formation and its ramification in our vasculatures, airways and lungs generate the pulmonary-vascular network to efficiently supply oxygen to the whole body. The airway tubes allow the airflow to alveolar structures, where blood cells inside the fine vascular tubes exchange carbon dioxide with oxygen to deliver it to the body (
Herriges and Morrisey 2014
;
Potente and Makinen 2017
;
Kishimoto and Morimoto 2021
). Surprisingly, a similar pulmonary-blood cell connection is also found in the fruit fly
Drosophila
. There, reiterative ramification of the epithelial tubes (the tracheal system) allows the airflow inside the body that directly delivers oxygen to the target cells (
Manning and Krasnow 1993
;
Samakovlis et al. 1996a
;
Hu and Castelli-Gair 1999
;
Ghabrial et al. 2003
), while the blood cells in the open circulatory system transiently attach to the fine airways to boost oxygen transport (
Shin et al. 2024
).
The lungs and the
Drosophila
airways both derive from 2D- primordial cell fields (
Romero et al. 2025
). Lung formation initiates as tube evaginations from the foregut epithelium (
Herriges and Morrisey 2014
;
Kishimoto and Morimoto 2021
). Distal ramification of these buds and further differentiation into distal airways and alveoli involve FGFR activation by extrinsic FGFs (
Min et al. 1998
;
Weinstein et al. 1998
;
Arman et al. 1999
;
Sekine et al. 1999
;
Brownfield et al. 2022
;
Jones et al. 2022
;
Sountoulidis et al. 2023
). Similarly,
Drosophila
airway formation initiates from cell primordia specified on the ectodermal plane, each invaginating to form a primitive cavity (
Perrimon et al. 1991
;
Manning and Krasnow 1993
;
Samakovlis et al. 1996a
;
Hu and Castelli-Gair 1999
). Extrinsic FGF/Branchless (Bnl) activates FGFR/Breathless (Btl) to guide branch ramification and network connection (
Klambt et al. 1992
;
Guillemin et al. 1996
;
Samakovlis et al. 1996a
;
Samakovlis et al. 1996b
;
Sutherland et al. 1996
) (
Figure 1A
). Accordingly, highly ramified fine tubes are generated around each branch terminus that serve for oxygen transport.
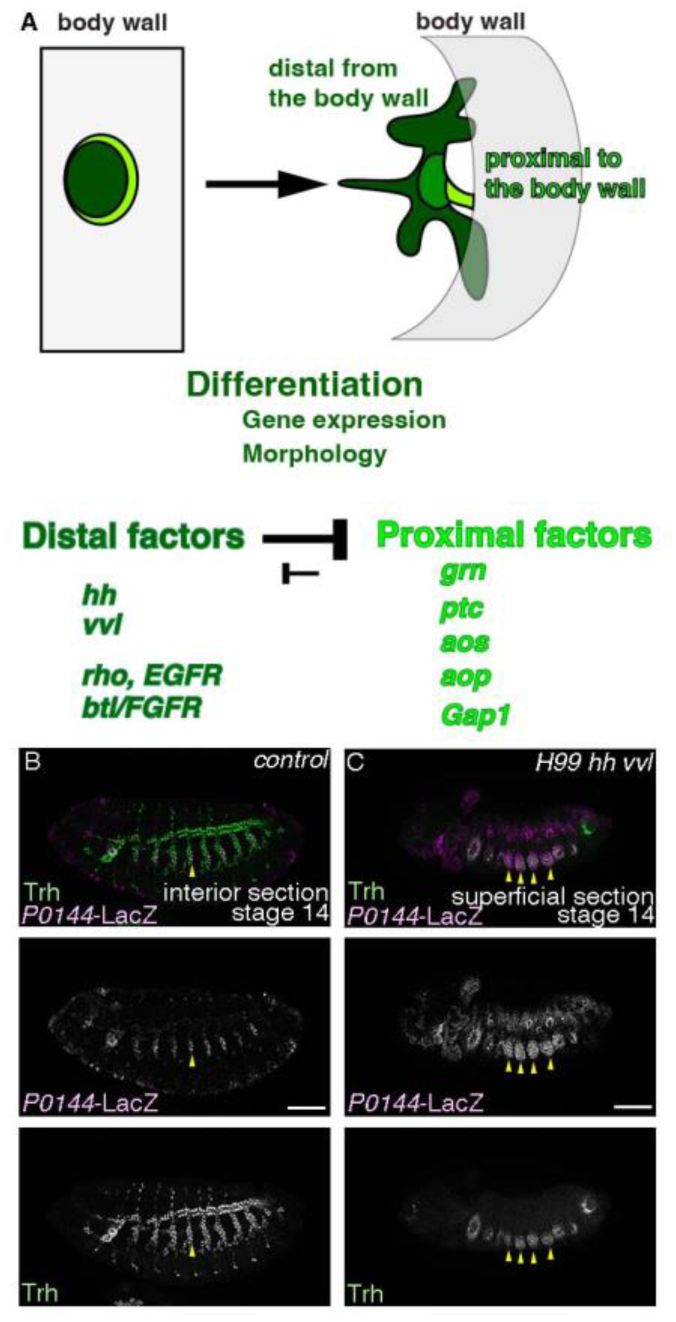
The distalizing factors
hh
and
vvl
dominantly promote invagination of the airway progenitors.
(A) A schema showing that invagination transforms the centro-peripheral patterning of the primordium into the proximo-distal patterning of the tubes. Extrinsic Hh and intrinsic Vvl dominantly promote the distal gene expression program (dark green domain) whereas
rho
and
btl/FGFR
are the distal factors that are known to distalize differentiation along both gene expression and morphology. The proximal factors including
grn
promote the proximal gene expression program (light green domain). (B-C) Single sections of lateral views of stage 14 embryos stained with Trh (a pan airway marker) and
P0144
-LacZ (a proximal marker). In the control (B), both the distal as well as the proximal airway progenitors (yellow arrowheads) invaginate to form a tube network whereas in
H99 hh vvl
mutants (
H99 hh
13C
vvl
utH599
P0144
homozygotes) (C), Trh
+
cells express the proximal marker
P0144
-LacZ and many of them stay at the epidermal layer (yellow arrowheads). Scale bar is 50 μm
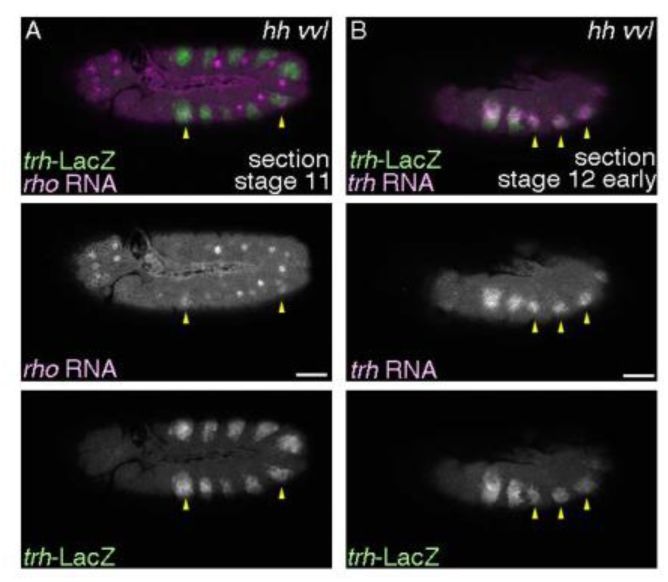
Expression of
rho
and
trh
in
hh vvl
double mutants,
hh
AC
vvl
utH599
/hh
13C
Df(vvl)
transheterozygotes.
(A) Weak and sporadic expression of
rho
is detected at early stage 11 in
hh vvl
double mutants (yellow arrowheads). (B) The airway progenitors frequently fail invagination in
hh vvl
double mutants as well and those cells retain
trh
expression at stage 12 (yellow arrowheads). Scale bar is 50 μm
trh
is the earliest TF gene marking the
Drosophila
airway primordia and the mature airways (
Isaac and Andrew 1996
;
Wilk et al. 1996
).
trh
has been regarded as the master TF of the
Drosophila
airways because tubes as well as differentiation markers are not detected in
trh
mutants at later stages (
Isaac and Andrew 1996
;
Wilk et al. 1996
;
Brodu and Casanova 2006
;
Sotillos et al. 2010
;
Chung et al. 2011
;
Matsuda et al. 2015b
). However, in
trh
null mutants, expression of
rhomboid
(
rho
) (
Bier et al. 1990
), a protease activating the EGFR ligand Spitz (
Schweitzer et al. 1995
) initiates in the primordia (
Ogura et al. 2018
) but its propagation (
Matsuda et al. 2015b
) fails (
Ogura et al. 2018
). Consistent with the role of
rho
in promoting primordial invagination (
Brodu and Casanova 2006
;
Nishimura et al. 2007
),
trh
mutants initiate primordia invagination but neither sustained invagination nor maintained tube structures (
Kondo and Hayashi 2019
). The initiation of invagination and the early localized
rho
expression in
trh
mutants suggest additional regulators, other than
trh
that are critically responsible for gene expression in airway primordia and for tubulogenesis.
We had previously shown that the proximo-distal axis of the
Drosophila
airway tubes is generated from the centro-peripheral patterning of the 2D primordia (
Matsuda et al. 2015b
). One set of genes, the distalizing factors,
hh
,
vvl
,
rho
and
btl/FGFR
cooperate to realize the distal gene expression program at the expense of the proximal one, whereas another set, the proximal factors including
grn
, negative regulators of
EGFR
signaling and
hh
signaling realize the proximal gene expression program (
Figure 1A
). The two programs establish distinct proximal and distal cellular domains in the branching network (
Matsuda et al. 2015b
). Two of the distalizing factors,
rho
and
btl/FGFR
promote primordia invagination but are not essential for tubulogenesis (
Brodu and Casanova 2006
;
Matsuda et al. 2015b
), indicating that essential regulators of primordial morphogenesis have been elusive.
Here, we report that in the absence of the remaining two distalizing factors
hh
and
vvl
, the trachea primodia cells frequently fail invagination and are only detected on the 2D plane. Therefore, extrinsic Hh and intrinsic Vvl represent the missing factors that cooperatively promote primordia invagination. Although
trh
,
vvl
or
grn
alone cannot define the airway progenitors, we propose that combinations of the three TFs can intrinsically define the airway progenitors, considering their subsequent roles on gene expression and morphogenesis. Even in the simple
Drosophila
airways, combinations of multiple factors induce the organ progenitor field and subsequent tubulogenesis to cope with physiological stress of respiration in a terrestrial environment.
Results and Discussion
The distalizing factors
hh
and
vvl
drive airway primordia invagination independent of
trh
Among the distalizing factors,
hh
,
vvl
,
rho
and
btl/FGFR
(
Figure 1A
),
hh vvl
double mutants show more extensive distal-to-proximal gene expression conversion than the loss of both
rho
and
bnl/FGF-btl/FGFR
signaling (
Matsuda et al. 2015b
). As
EGFR
(also known as
torpedo/top
or
faint little ball)
and
btl/FGFR
signaling also orchestrate the distal morphogenetic processes, including primordial cell invagination (
Llimargas and Casanova 1999
;
Brodu and Casanova 2006
;
Nishimura et al. 2007
;
Kondo and Hayashi 2013
) and subsequent branching (
Glazer and Shilo 1991
;
Klambt et al. 1992
;
Sutherland et al. 1996
;
Llimargas and Casanova 1999
;
Matsuda et al. 2015b
),
hh
and
vvl
are expected to control the distal morphogenetic program as well. In the absence of
hh
and
vvl
,
rho
expression is detected at early stages sporadically and weakly (
Figure 1-figure supplement 1A
) and
btl
expression initiates in the primordia, but both fade away shortly thereafter (
Matsuda et al. 2015b
). Despite the initial expression of
rho
and
btl/FGFR
in
hh vvl
mutants,
hh vvl
embryos show severer branching phenotypes than the
rho btl
mutants (
Matsuda et al. 2015b
).
We thus investigated potential invagination defects of
trh
positive cells in
hh vvl
double mutants and detected aberrant expression of both
trh
RNA and
trh
-LacZ persisting on the epidermal plane at stage 12 (
Figure 1-figure supplement 1B
). However, massive ectodermal apoptosis precluded analysis at later stages. In
H99 hh vvl
triple mutants, where apoptosis is suppressed by deletion of major pro-apoptotic genes (
White et al. 1994
), we often detected failure in invagination, where all the Trh+ cells remained in a 2D plane at stages 12-13 (
Figure 1A-B
). By stage 15, small cavities are detected in most metameres of
H99 hh vvl
mutants, suggesting that proximal cell types invaginate by a mechanism independent of distal cell types differentiation. Taken together, the two distalizing factors
hh
and
vvl
define the central/distal cell identity, controlling both gene expression and cell invagination. It follows that 3 intrinsic TFs, Trh, Vvl and Grn cover the major functional aspects of airway progenitor differentiation.
dpp/BMP
specifies the airway progenitors along the D-V axis
We further investigated upstream determinants of the primordial field defined by the expression of these 3 TFs. We first allocated their expression domains along the D-V axis of the embryo. The surface of the embryonic trunk at mid embryogenesis is largely divided into 3 sectors along the D-V axis, amnioserosa, the dorsal and the ventral ectoderm (
Figure 2A
) (
Wharton et al. 1993
;
von Ohlen and Doe 2000
). Both the dorsal and the ventral ectoderm are further subdivided into 3 parts along the D-V axis, medial, intermediate and lateral (
Figure 2A
) (
von Ohlen and Doe 2000
). The dorsal intermediate column marked with
caupolican (caup)/araucan (ara)
spans several cells located dorsally to the
trh
-LacZ positive spiracular branches (
Figure 2-figure supplement 1A
), whereas the Grn-GFP expression domain, which also marks the most proximal spiracular branches extends ventrally from the dorso-lateral ectoderm (
Figure 2- figure supplement 1B
). On the other hand, the ventral limits of the initial
trh
expression at stage 10 abut
Dichaete
-LacZ (
D
-LacZ) expressing cells in the intermediate and medial columns of the ventral ectoderm (
Figure 2B-C
) (
Zhao and Skeath 2002
). Thus, the initial
trh
expression straddles from the ventro-lateral column to halfway to the dorso-lateral column.
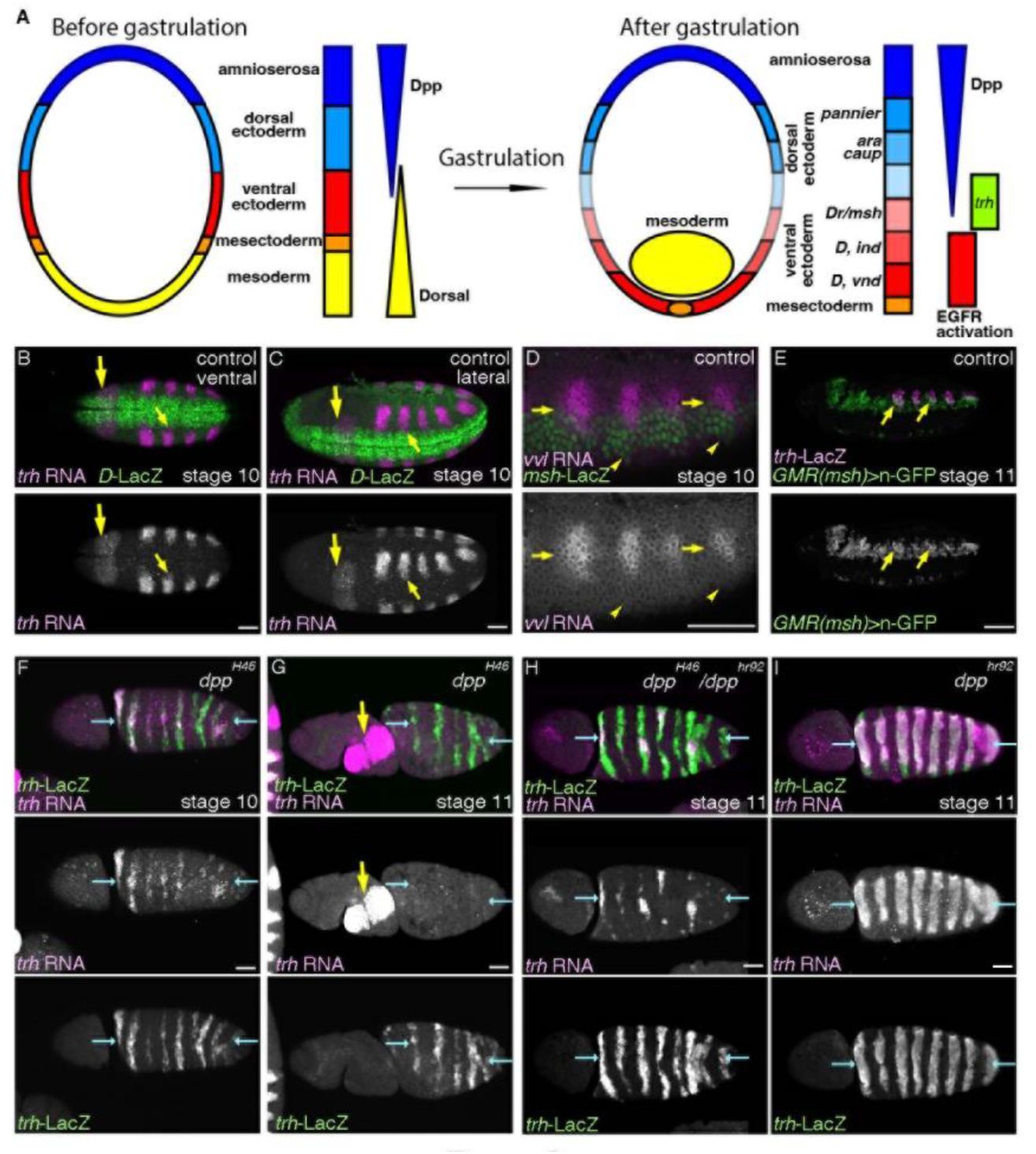
Medium Dpp/BMP activity promotes
trh
expression and the airway progenitors.
(A) A schema showing that the embryonic trunk is divided into discrete domains along the D-V axis. Out of the 5 domains (amnioserosa, dorsal ectoderm, ventral ectoderm, mesectoderm and mesoderm), mesoderm invaginates during gastrulation. Graded activities of a TF
dorsal
orchestrates the ventral domains whereas Dpp/BMP signaling orchestrates the dorsal domains. Differential expression of TFs subdivides both the dorsal and the ventral ectoderm into medial, intermediate and lateral columns. EGFR is dynamically activated to establish the ventro-intermediate column (
Yagi et al. 1998
). Expression of TFs marks the column of dorso-medial (
pannier
), dorso-intermediate (
araucan/ara, caupolican/caup
), ventrol-lateral (
Dr/msh
), ventro-intermediate (
Dicheaete/D, intermediate neuroblasts defective/ind
), ventro-medial (
D, ventral nervous system defective/vnd
). (B-E) Distribution of
trh
and
vvl
transcripts relative to the ectodermal subdivision along the D-V axis. (B-C) Ventral and lateral views stage 10 embryos. The ventral limit of
trh
expression abuts the dorsal limit of
D
-LacZ expression (small yellow arrows) which marks the ventro-medial and the ventro-intermediate columns. Large arrows show
trh
expression in the salivary gland primordia. (D-E)
vvl
expression at stage 10 straddles the border between the dorsal ectoderm and the
Dr/msh
-LacZ positive ventro-lateral ectoderm (D, yellow arrows) whereas the ventral limit of
vvl
expression is some cells away from the ventral limit of
Dr/msh
-LacZ expression (D, yellow arrowheads). An enhancer fragment of
Dr/msh
active in the ventro-lateral column marks the ventral parts of the airway tubes at stage 11 (E, yellow arrows). (F-I) Expression of trh-LacZ and trh transcripts in allelic series of dpp mutants. Dorsal views of embryos where the presumptive dorsal midline are marked with blue arrows. In
dpp
null mutants (F, G),
trh
expression detected with
trh
-LacZ or
trh
RNA expands to the dorsal midline (blue arrows). However at later stages,
trh
RNA is not maintained though
trh
-LacZ positive cells remain (G). Note that in dpp null mutants the body is twisted. In the milder condition (H),
trh
RNA is sporadically maintained near the dorsal midline whereas in
dpp
hypomorph homozygotes (I),
trh
RNA is detected in many of the expanded progenitor areas. Scale bar is 50 μm
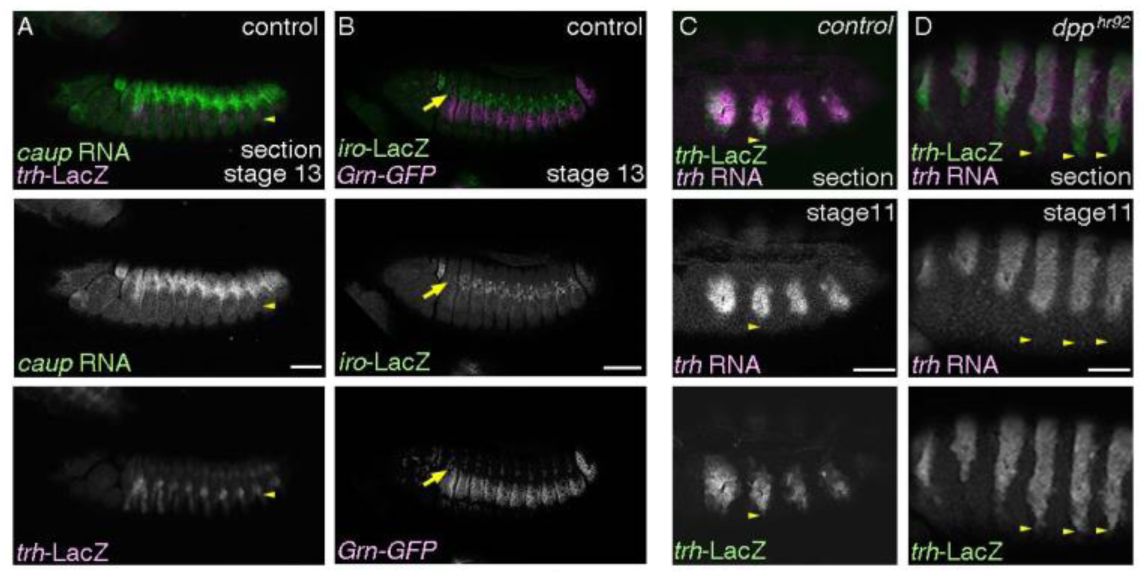
The proximal airways and the ectodermal subdivision.
(A-B) Lateral views of stage 13 embryos. The spiracular branch cells (
trh
-LacZ, yellow arrowheads) are several cells ventrally to the
caup
RNA expressing cells (A). Grn-GFP expresses not only in the spiracular branches but also in the dorso-lateral ectodermal cells that reside ventrally to the
iroquois
-LacZ (
iro
-LacZ)/
araucan
-LacZ) positive dorso-intermediate column (B, yellow arrows). (C-D) Lateral views of stage 11 embryos.
trh
-LacZ positive cells that lose
trh
RNA are pronounced in the ventral parts of the control (C). This becomes more pronounced in the
dpp
hypomorhps (D). Yellow arrowheads mark the ventral limits of
trh
-LacZ expression. A, C and D are single sections. Scale bar is 50 μm
Compared to
trh
expression,
vvl
expression in the airway primordia is more restricted (
Figure 2D
).
vvl
expression straddles the boundary of the dorsal and the ventral ectoderm demarcated by
Drop (Dr)/muscle homeobox (msh)
whereas the ventral limit of
vvl
expression is far from the ventro-intermediate column (
Figure 2D
) (
von Ohlen and Doe 2000
). Consistently, the ventro-lateral ectoderm enhancer of
Dr/msh
(
Pfeiffer et al. 2008
;
Manning et al. 2012
) marks the ventral parts of the invaginated airway progenitors (
Figure 2E
).
Medium level of Dpp/BMP signaling positively regulates the expression of
vvl
and
grn
(
Matsuda et al. 2015b
) but its function of
trh
regulation is only partially investigated. In the absence of Dpp/BMP, the dorsal 2 sectors, amnioserosa and the dorsal ectoderm take the cell differentiation program of the ventro-lateral ectoderm (
von Ohlen and Doe 2000
). Consistent with that, the initial
trh
expression straddles the ventro-lateral ectoderm (
Figure 2B-C
),
trh
expression expands to the dorsal midline in
dpp
null mutants (
Figure 2F
) (
Isaac and Andrew 1996
). However,
trh
expression is extinguished by stage 12 in
dpp
null mutants (
Figure 2G
), consistent with the Dpp/BMP’s role on expression of
vvl
and
grn
, which in turn maintain
trh
expression (
Matsuda et al. 2015b
). In milder inactivation conditions of
dpp/BMP
hypomorphic mutants (
dpp
H46
/dpp
hr92
trans-heterozygotes) (
Wharton et al. 1993
),
trh
maintenance, if any, occurs only around the dorsal midline, where reduced Dpp/BMP activity levels are presumed to be sporadically present (
Figure 2H
). In even milder conditions of
dpp/BMP
inactivation in
dpp
hr92
hypomorphs (
Wharton et al. 1993
),
trh
maintenance occurs in the more ventral cells as well (
Figure 2I
). However, we note that concomitant with reduction of Dpp/BMP activities, more cells in the ventral part of the initial trh expression domain fail to maintain
trh
expression (
Figure 2F-I
,
Figure 2-figure supplement 1C-D
). Loss of the ventral
trh
expression appears to occur also in wild type embryos (
Figure 2-figure supplement 1C
), possibly reflecting that the most ventral cells are farthest from the Dpp/BMP source.
The distal and the proximal progenitors are specified in
CycA
mutants (
Matsuda et al. 2015b
), where the last progenitor mitosis does not occur (
Beitel and Krasnow 2000
). As the distal cells are double positive for
trh
and
vvl
and occupy 90% of the mature airways, we speculate that Trh
+
cells that are in and near the Vvl
+
areas are the airway progenitors whereas the remaining Trh
+
cells would become epidermal. These airway progenitors are specified straddling the canonical dorsal-ventral ectoderm boundary. We conclude that an optimal Dpp/BMP activity specifies the airway progenitors along the D-V axis.
The radial EGFR signaling primes the airway progenitors and realizes airway differentiation along the P-D axis
Airway cells expressing late differentiation markers are reduced in number in
H99 EGFR btl/FGFR
mutants, where anti-apoptotic functions of
btl/FGFR
and
EGFR
are compensated by
H99
deficiency (
Matsuda et al. 2015b
). These missing cells prompted us to investigate the potential roles of the two RTKs on
trh
expression earlier.
trh
expression is comparable in the distal and the proximal cells at stage 12 in
H99 btl/FGFR
mutants (
Figure 3A
). In contrast,
trh
expression in the proximal area becomes reduced or extinguished at stage 12 in
H99 EGFR
mutants (
Figure 3B
). Moreover,
trh
expression in the main airways is very much reduced in
H99 EGFR btl/FGFR
mutants, leaving only residual expression in a subset of the invaginated cells (
Figure 3C
). Thus, EGFR signaling is the predominant factor promoting maintenance of
trh
expression. In its absence, this function can be compensated by Btl/FGFR signaling (
Figure 3D
).
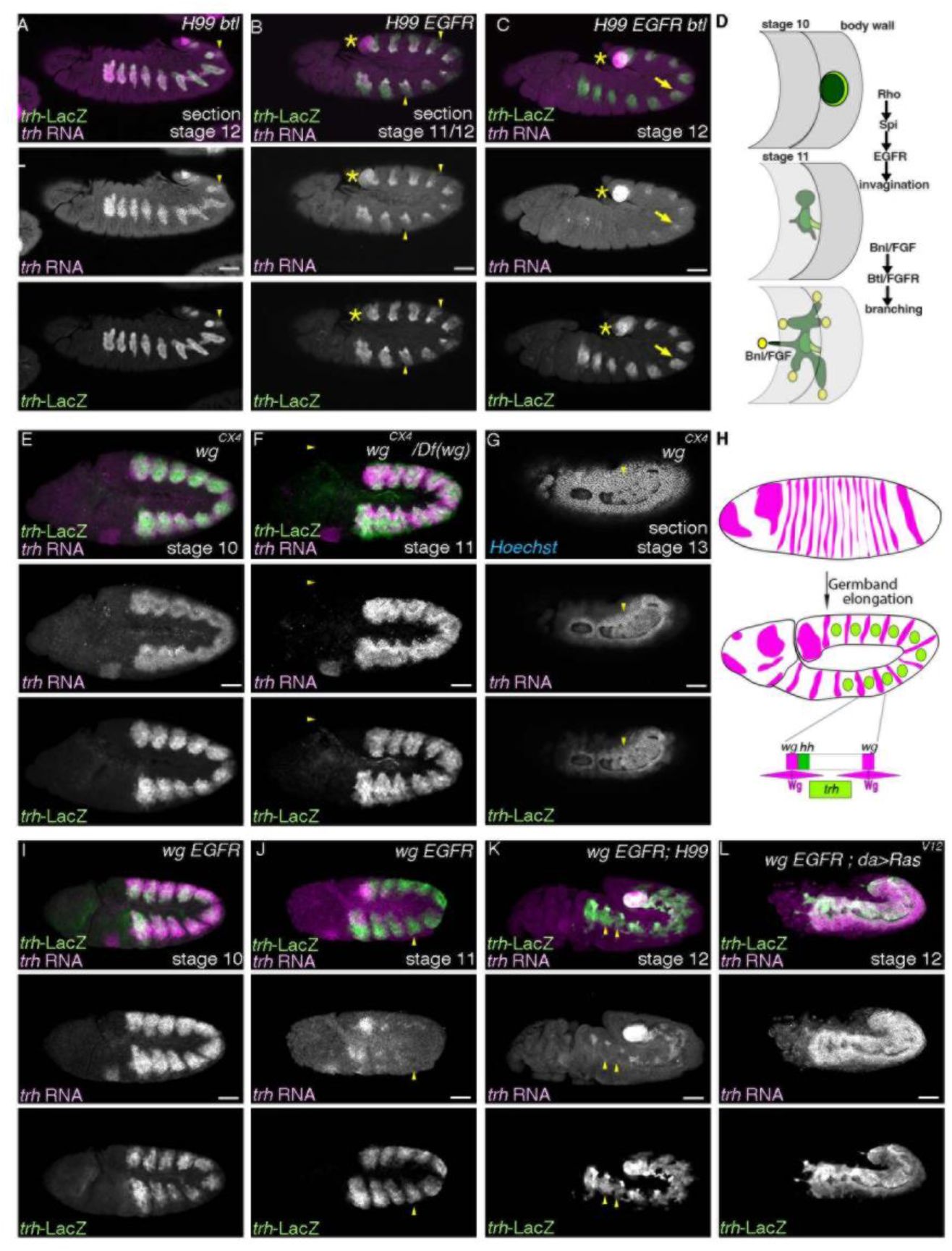
Maintenance of
trh
expression does not follow changes in tissue architecture Lateral views of embryos stained with
trh
-LacZ and
trh
transcripts.
(A-D) In
H99 btl/FGFR
mutants (
H99 btl
Δoh10
/H99 btl
Δoh24-1
) (A),
trh
expression is detected both in the distal and the proximal regions whereas in
H99 EGFR
mutants (
top
f2
/top
f24
; H99
) (B),
trh
expression in the proximal regions is significantly reduced (yellow arrowheads). In
H99 EGFR btl/FGFR
mutants (
top
f2
/top
f24
; H99 btl
Δoh10
/H99 btl
Δoh24-1
) (C), residual
trh
expression is detected in parts of the invaginated cells (yellow arrows). Asterisks mark
trh
expression in the posterior spiracle primordia. A schema in D shows stages and functions of RTK activation in the airway progenitors. (E-L) In the absence of Wg/WNT (E-G), which is expressed in stripes along the A-P axis (H), the airway progenitor areas expand along the A-P axis.
trh
-LacZ and
trh
RNA are largely co-expressed since before invagination. Arrowheads in G show that
trh
is expressed in cells that take the 2D planar configuration. In
EGFR wg
double mutants (
top
f24
wg
CX4
homozygotes) (I, J), maintenance of
trh
RNA becomes defective at around the stage of invagination (compare I and J).
trh
maintenance is restored not by suppression of apoptosis (
top
f24
wg
CX4
;H99
) (K) but by
daughterless (da)
-Gal4 driven overexpression of Ras
V12
(
top
f24
wg
CX4
;da-Gal4/UAS-Ras
V12
) (L). Note that
trh
RNA is not detected in cells positive for
trh
-LacZ in J and K (yellow arrowheads). Scale bar is 50 μm

trh
expression in
aos Gap1
double mutants do not respect tissue architecture.
(A-B) Lateral views of
aos Gap1
mutant embryo stained with
trh
-LacZ and
trh
transcripts. A projection (A) or a single section (B).
trh
expression is often detected in planes at the embryo surface (yellow arrows). Scale bar is 50 μm
In contrast to a model, where maintenance of
trh
expression correlates with the transition of primordial cells from a planar 2D into a 3D tubular tissue architecture (
Kondo and Hayashi 2019
), we show that
trh
expression is maintained in the cells on the 2D ectodermal plane in the
hh vvl
double mutants (
Figure 1C
,
Figure 1-figure supplement 1B
). Additionally,
trh
expression is very much reduced in the invaginated cells of
H99 EGFR btl/FGFR
mutants (
Figure 3C
), uncoupling
trh
gene expression from the morphogenetic process of invagination.
To further test if tissue architecture contributes to the maintenance of
trh
expression, we made use of mutations in
wingless (wg)/WNT
, which is expressed in longitudinal stripes and represses expression of
trh
and
vvl
along the anterior-posterior (A-P) axis (
Figure 3E-H
) (
de Celis et al. 1995
;
Wilk et al. 1996
). In
wg
mutants, some of the progenitors fail to internalize so that cell expressing either distal or proximal markers are detected at the 2D planar embryo surface (
Oda et al. 1994
;
Matsuda et al. 2015b
). Correspondingly,
trh
RNA and
trh-
LacZ are detected on the embryo surface in
wg
mutants at stage 13 and later (
Figure 3G
).
Similar to
wg
mutants, the initial
trh
expression expands along the A-P axis in
wg EGFR
double mutants (
Figure 3I
). However,
trh
RNA expression becomes very weak when most cells still reside on the 2D planes around the stage of primordia invagination (
Figure 3J
). This significant reduction of
trh
expression domain is still evident even in
H99 wg EGFR
mutants where
H99
deficiency suppresses apoptosis (
Figure 3K
). Together, we conclude that tissue architecture is dispensable for
trh
maintenance.
We suggest that
trh
maintenance is stimulated concurrently with the robust EGFR activation that occurs and propagates in the 2D airway primordia (
Gabay et al. 1997a
;
Gabay et al. 1997b
;
Wappner et al. 1997
;
Matsuda et al. 2015b
). EGFR activation also distalizes gene expression (
Matsuda et al. 2015b
) and initiates morphogenesis (
Brodu and Casanova 2006
;
Nishimura et al. 2007
). In this scenario, rather than indirectly through tubulogenesis (
Kondo and Hayashi 2019
), RTK signaling directly promotes
trh
maintenance irrespective of tissue geometry. We note that in double mutants of
aos
and
Gap1
, which are both negative regulators of EGFR signaling, there is expression of airway distal marker on the embryo surface (
Matsuda et al. 2015b
). This is accompanied by detection of
trh
expression on the embryo surface (Figure 3-figure supplement 3A-B).
Thus, subsequent to the Dpp/BMP mediated specification of the airway progenitors along the D-V axis, the radial EGFR signaling sustains
trh
expression in the progenitors and initiates airway cell differentiation along the P-D axis.
DRaf/MAPKKK and Dsor1/MAPKK are required for EGFR mediated priming of the airway progenitors
Trh activity can be boosted by the PI3kinase-PKB pathway through phosphorylation of Serine 665 (
Jin et al. 2001
). However, it is not clear how far this pathway is required
in vivo
for Trh activity and
trh
autoregulation. We thus investigated if EGFR mediated priming and
trh
expression maintenance in the airway progenitors require the Ras-MAPK pathway (
Figure 4A
) (
Hou et al. 1995
;
Mishra et al. 2005
). When MAPKK-kinase/Draf or MAPK-kinase/Dsor1 is depleted from the embryos with the
ovo-FRT
germline clone technique (
Hou et al. 1995
;
Chou and Perrimon 1996
),
trh
expression initiates, as detected with
trh
-LacZ enhancer trap (
Figure 4B-C
). However, around the stage of primordial invagination,
trh
expression is very much reduced, in both non-invaginated or invaginated cells (
Figure 4B-C
).
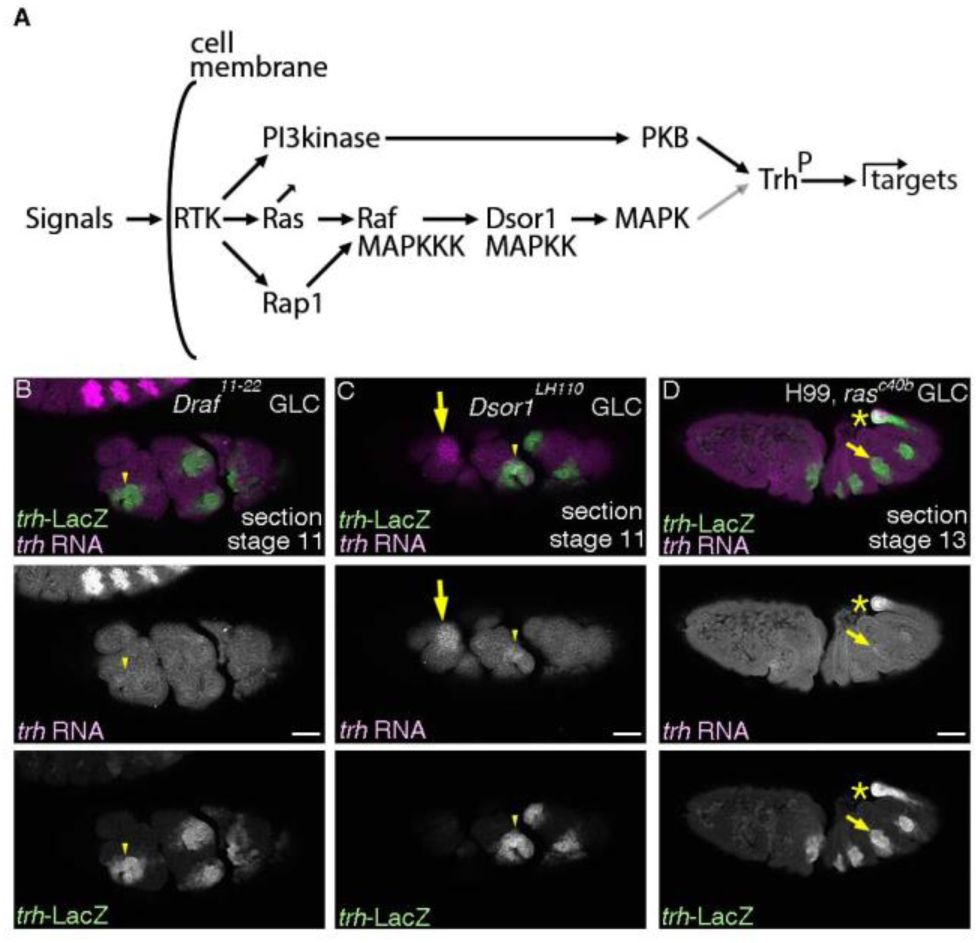
RTKs require the Ras-Raf/MAPKKK-Dsor1/MAPKK pathway for maintenance of
trh
expression.
(A) A schema showing that RTK activation involves several downstream signaling branches including the Ras-MAPK pathway and the PI3kinase-PKB pathway. PKB is known to phosphorylate Trh and to upregulate Trh transcriptional activity (black arrow). MAPK may act the same (gray arrow). (B-D) Lateral section views of embryos stained with
trh
-LacZ and
trh
transcripts. In the complete absence of Draf/MAPKKK (B) or Dsor1/MAPKK (C),
trh
RNA is hardly maintained in the cells positive for
trh
-LacZ, either invaginated or non-invaginated. In the complete absence of
Drosophila Ras1/Ras85D
, introducing
H99
deficiency suppressing apoptosis allows visualization of airway development till later stages (D), demonstrating residual
trh
expression in parts of the invaginated cells. Yellow arrows in C show
trh
expression in the salivary gland. Yellow asterisks in D mark
trh
expression in the posterior spiracle. Scale bar is 50 μm
Ras
V12
boosts Trh activity
Ras
(
Ras85D
or
Ras1
in
Drosophila
) is one of the major signaling mediators of RTKs through Draf/MAPKKK-Dsor1/MAPKK-MAPK (
Hou et al. 1995
;
Mishra et al. 2005
) and through Pi3kinase- PKB (
Figure 4A
) (
Orme et al. 2006
). Consistent with the major role of
Ras85D/Ras1
in RTK signaling, comparable defects in maintenance of
trh
expression are detected in
EGFR btl/FGFR
double mutants and upon complete loss of
Ras85D/Ras1
, when anti-apoptotic functions are compensated by the
H99
chromosomal deletion (
Figure 3C
,
4D
). Conversely,
trh
maintenance in
EGFR wg/WNT
double mutants is significantly rescued resembling
wg/WNT
single mutants alone upon overexpression of the gain of function form of Ras85D, Ras
V12
(
Fortini et al. 1992
) (
Figure 3L
). Thus,
Ras
serves as the major signaling transducer to promote
trh
maintenance downstream of EGFR and Btl/FGFR.
The mechanisms by which,
Ras
promotes
trh
expression may involve boosting Trh activity because
trh
auto-regulates its own expression (
Figure 4A
) (
Wilk et al. 1996
;
Sotillos et al. 2010
;
Chung et al. 2011
). Boosting of Trh activity may involve PKB mediated phosphorylation of Trh (
Jin et al. 2001
). Alternatively, inferred from the positive roles of MAPK mediated phosphorylation of Hif1, a
trh
homologue (
Mylonis et al. 2006
), MAPK may directly upregulate Trh activity. Consistently, overexpression of the phosphomimetic form of Trh, Trh
S665D
or simultaneous double overexpression of Ras
V12
and Trh
WT
significantly induced ectopic expression of
trh
-LacZ, whereas single overexpression of Ras
V12
or Trh
WT
did not (
Figure 5C
). The Ras
V12
induced increase of Trh activity is also detected on the expression of both the distal (
btl
and Gasp/2A12) and the proximal marker (
upd
and
P0144
-LacZ) as well (
Figure 5A-F
). We conclude that Ras activation downstream of RTKs is sufficient to upregulate Trh activity toward its downstream targets (
Figure 4A
).
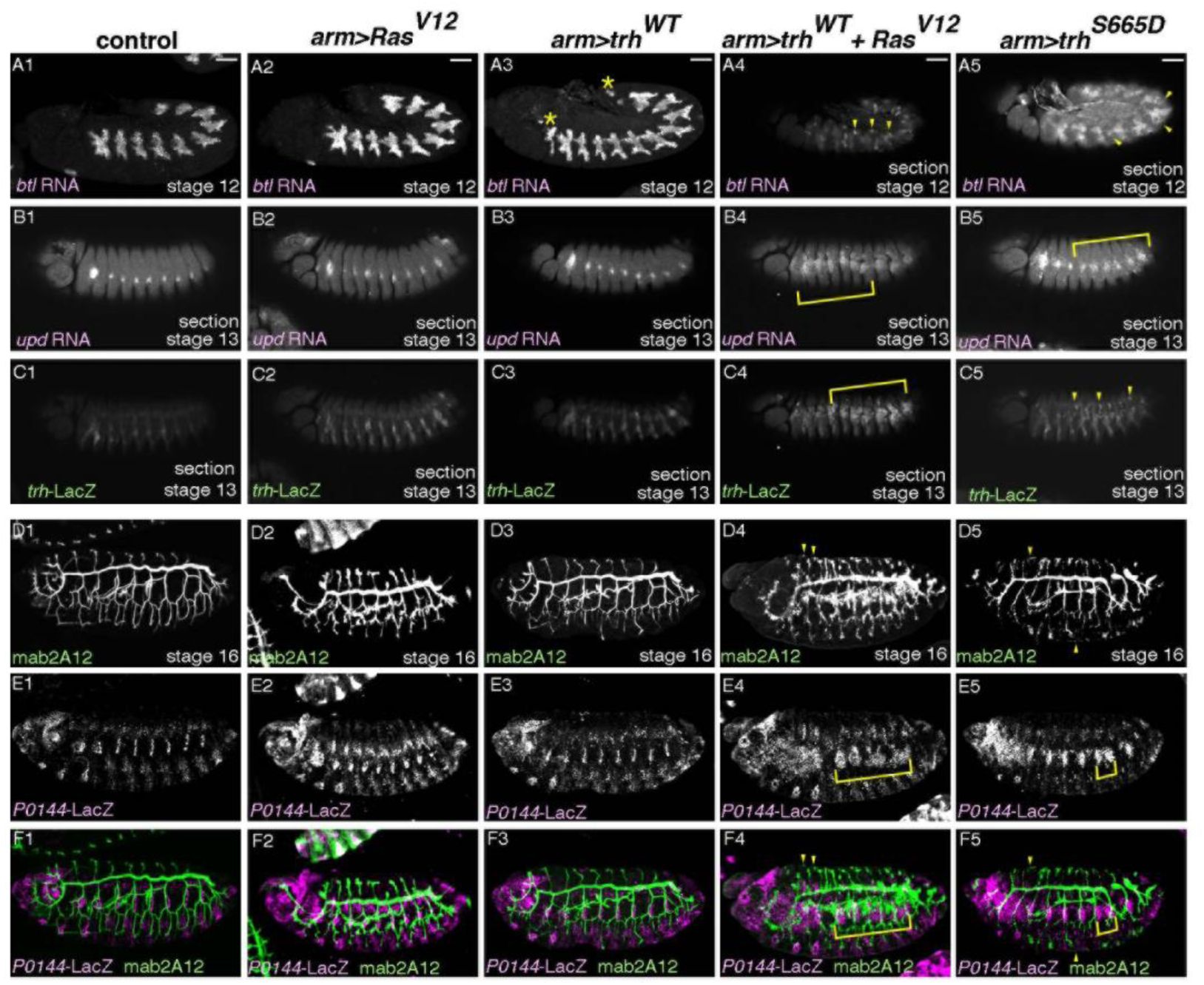
Ras
V12
boosts Trh activity toward downstream genes.
(A-F) Lateral views of embryos stained as indicated. Compared to the control (first column), single overexpression of Ras
V12
(second column) or Trh
WT
(third column), simultaneous overexpression of Ras
V12
and Trh
WT
(fourth column) or single overexpression of Trh
S665D
(fifth column) induces ectopic expression of
btl
(A),
upd
(B),
trh
-LacZ (C), mab2A12 (Gasp) (D, F) or
P0144
-LacZ (E, F), which are marked by yellow arrowhead or yellow blankets. Asterisks in A3 marks ectopic progenitors in the anterior and the posterior segments. Scale bar is 50 μm.
Multiple redundant regulators cooperate during early Drosophila airway tubulogenesis
Tubes are common structures in multicellular organisms and tubular organs fulfill essential functions for life (
Romero et al. 2025
). An active field in stem cell biology engages in artificial organ generation
in vitro
to complement tubular organ dysfunction (
Lancaster and Knoblich 2014
). In
vivo
studies of tubulogenesis may aid such
in vitro
organoid development. In line with the revised view that a simple master regulator does not dictate all the steps of the
Drosophila
airway differentiation (
de Celis et al. 1995
;
Llimargas and Casanova 1997
;
Chen et al. 1998
;
Boube et al. 2000
;
Matsuda et al. 2015b
;
Ogura et al. 2018
;
Kondo and Hayashi 2019
), we characterize that the cooperation of both extrinsic and intrinsic, partly-redundant regulators ensures their robust morphogenesis.
The airway progenitor specification and their subsequent differentiation is associated with continuous expression of
trh
(
Isaac and Andrew 1996
;
Wilk et al. 1996
).
trh
is necessary for expression of all airway differentiation markers and for establishment of the airway tubes (
Isaac and Andrew 1996
;
Wilk et al. 1996
;
Sotillos et al. 2010
;
Chung et al. 2011
;
Matsuda et al. 2015b
), assigning it as a master TF of
Drosophila
airway tubulogenesis. Surprisingly, however,
trh
mutant cells invaginate to form tubes, but later revert to a planar configuration (
Kondo and Hayashi 2019
). Resolving the issue of what promotes progenitor invagination other than Trh, we identified extrinsic Hh and intrinsic Vvl as the dominant regulators of invagination, leaving
trh
expression intact.
The airway progenitors are intrinsically defined with at least 3 TFs,
trh
,
vvl
and
grn
.
vvl
defines the central/distal progenitors that invaginate first, whereas
grn
defines the peripheral/proximal progenitors that invaginate later, reflecting the proximo-distal differences of the airways (
Matsuda et al. 2015b
). Maintenance of
trh
expression is promoted significantly by auto-regulation (
Wilk et al. 1996
;
Sotillos et al. 2010
;
Chung et al. 2011
). However, regulation of
trh
expression is far more complex, requiring discrete regulations along the A-P, the D-V and the radial/proximo-distal (P-D) axis (
Figure 6
).
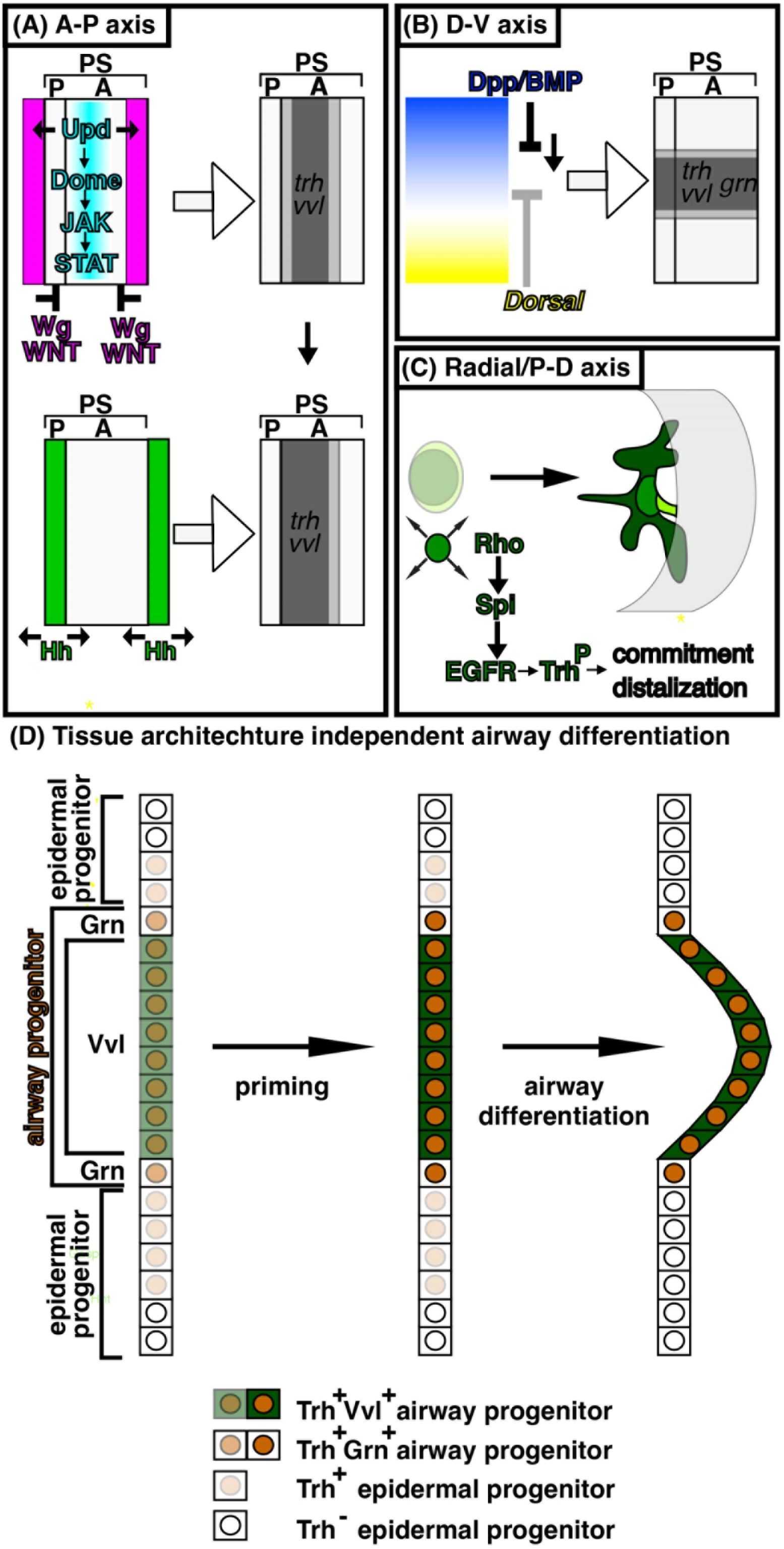
A model for specification and priming of the airway progenitors along the three body axes Inputs along the A-P axis and the D-V axis generate radial patterning of the airway progenitors to realize the proximo-distal differences of gene expression and morphology.
(A) A-P axis; Segments and para-segments (PS) are units of
Drosophila
ectoderm segmentation along the A-P axis. One PS is composed of the posterior of a segment and the anterior of its posterior neighboring segment. Wg/WNT (coloured pink) represses
trh
and
vvl
whereas Upd ligands signal through Domeless-JAK-STAT (coloured blue) to induce them, which together generate a graded airway progenitor field along the A-P axis (coloured gray). The
upd
expression domain is a guess.
upd
expression is dynamic (
Harrison et al. 1998
;
Sotillos et al. 2010
) and it is not known which
upd
expression is required for inducing the airway primordia.
trh
expression initiates already at stage 8 (
Isaac and Andrew 1996
) whereas
vvl
expression starts at stage 10. Hh (coloured green) positively regulates expression of
vvl
, thereby distalizes the progenitor field. (B) D-V axis; The ventrally active TF Dorsal (coloured yellow) restricts
dpp/BMP
expression to the dorsal domain. Dpp/BMP (coloured blue) in turn orchestrates the dorsal parts of the embryos. Midium Dpp/BMP activities promote expression of
trh
,
vvl
and
grn
to generate a graded airway progenitor field along the D-V axis. (C) P-D axis; The radial patterning is realized by Rho mediated activation of EGFR. EGFR activation primes the airway progenitors to commit to the airway differentiation program. At the same time, EGFR distalizes the airway differentiation along both gene expression and morphology to establish the P-D axis of the invaginated tubes. Spreading of
rho
expression and promotion of
trh
maintenance may involve EGFR mediated activation of TFs like Trh and Vvl, which boosts their transcriptional activities toward downstream genes including
rho
and
trh
. (D) A model of tissue-architecture-independent regulation of trh expression.
trh
expressing cells (brown coloured nucleus) are composed of the epidermal progenitors (pale brown) and the airway progenitors (brown), the latter of which is further classified into
vvl
expressing central/distal cells (green cytoplasm) and
grn
expressing peripheral/proximal cells. Each progenitor type is specified on the 2D cell fields based on the cues along the A-P and the D-V axes (left). Radial EGFR signaling primes the airway progenitors (center, dark brown), to realize morphogenetic and transcriptional differentiation along the P-D axis (right).
trh
expression in the epidermal progenitors ceases based on the epidermal differentiation programs. Airway progenitor invagination may be aided by segregation of the 2 different cell types whereas invagination processes may finetune the cell type determination processes. Note that
grn
expression in the epidermal progenitors is omitted.
Along the D-V axis, our results show that intermediate Dpp/BMP activity promotes maintenance of
trh
expression whereas high level Dpp/BMP activity is known to be repressive on initiation of
trh
expression (
Isaac and Andrew 1996
;
Wilk et al. 1996
). Therefore, Dpp/BMP has two opposing effects on
trh
expression.
Along the A-P axis, segmentally repeated expression of Unpaired ligands and the resultant activation of Domeless-JAK-STAT signaling precedes and may set where expression of
trh
and
vvl
initiates (
Brown et al. 2001
;
Sotillos et al. 2010
). The airway field is repressed in the embryonic head and the tail by a zinc finger TF
spalt
(
Boube et al. 2000
) whereas it is segmentally repressed in the trunk by Wg/WNT (
de Celis et al. 1995
;
Wilk et al. 1996
). It is not known what initiates the remaining expression of
trh
and
vvl
in the absence of Unpaired family ligands-Domeless-JAK-STAT signaling (
Brown et al. 2001
).
Along the radial/P-D axis, the centro-peripheral spreading of Rho-mediated EGFR activation in the 2D progenitor fields promotes maintenance of
trh
expression. Co-expression of Trh and Ras
V12
potentiates ectopic Trh activity toward downstream genes. Therefore, we suggest that EGFR mediated boosting of TFs like Trh or Vvl may underly
trh
maintenance (
Wilk et al. 1996
) and the sequential spreading of
rho
expression (
Matsuda et al. 2015b
), generating a wave of feed-forward loops for progenitor priming and airway differentiation.
Also, we previously showed that
grn
potentiates
trh
maintenance in the peripheral/proximal cells whereas
vvl
circumvents the
trh
expression dependency on
grn
in the central/distal cells (
Matsuda et al. 2015b
), arguing for region-specific requirements for
trh
maintenance in the primordium.
In our model, it is pre-determined which cells in the
trh
expressing 2D fields contribute to airways or epidermis before invagination (
Figure 6D
). Cells not receiving proper amounts of positive inputs from like optimal Dpp/BMP activities or enough EGFR activities or that receive too much negative inputs like Wg/WNT, would cease
trh
transcription. The timing of transcriptional silencing and degradation of the remaining transcripts and the proteins would determine when the cells lose
trh
products. Recently, it was discovered that micro-peptides encoded by
polished rice/tarsal-less
promote
trh
expression by suppressing the repressor activity of a zinc finger TF Ovo/Shaven-baby (
Mizuno et al. 2026
). It is intriguing, where this circuit fits into the regulatory modes of
trh
expression we described above.
Multiple partially redundant systems on initiation and maintenance of gene expression and morphology sustain development of even simple organs like the
Drosophila
airways. The crucial roles of tubular organs for survival could be the driving force in evolving multiple overlying regulatory schemes, which may secure development of essential organs even when one regulatory scheme becomes dysfunctional.
Materials and methods
Fly genetics and histochemistry of embryos were done as previously described (
Matsuda et al. 2015a
;
Matsuda et al. 2015b
). Marker genes were typically introduced as a single copy, unless otherwise noted.
Fly strains used in this study are:
aos
Δ7
(BDSC 1004)
arm-Gal4
(BDSC 1560)
btl-Gal4
(a gift from Dr. S. Hayashi)
D-lacZ
(a gift from Dr. J. Nambu and Dr. S. Russel)
Draf
11-22
FRT101
(a gift from Dr. N. Perrimon)
Dsor1
LH110
FRT101
(a gift from Dr. N. Perrimon)
btl
Δoh10
(
Ohshiro and Saigo 1997
)
btl
Δoh24-1
(
Ohshiro and Saigo 1997
)
da-Gal4
(BDSC 8641)
dpp
H46
(BDSC 2061)
dpp
hr92
(BDSC 2069)
Dr/msh-lacZ
(a gift from Dr. A. Nose)
Df(2L)Exel6017=Df(wg)
(BDSC 7503)
Df(3L)Exel6109=Df(vvl)
(BDSC 7588)
Gap1
B2
(a gift from Dr. N. Perrimon)
GMR19D03
(BDSC 48846)
grn-GFP
(BDSC 58483)
H99=Df(3L)H99
(BDSC 1576)
hh
13C
(
Hosono et al. 2003
)
hh
AC
(BDSC 1749)
hs-FLP
(BDSC)
iroquois-lacZ
(a gift from Dr. S. Campuzano)
ovoD1 FRT101; hs-FLP
(BDSC 1813)
ovoD1 FRT82B
(BDSC 2149)
P0144-lacZ
(a gift from Dr. W. Janning, Flyview)
Ras85D
Δc40b
(a gift from Dr. N. Perrimon and Dr. C. A. Berg)
top
f2
(BDSC 2768)
top
f24
(a gift from Dr. K. Moses)
trh-lacZ=1-eve-1
(a gift from Dr. N. Perrimon)
UAS-GFP.nls
(BDSC 4776)
UAS-trh
WT
/CyO
(a gift from Dr. A. S. Manoukian)
UAS-trh
S665D
(a gift from Dr. A. S. Manoukian)
UAS-Ras
V12
/CyO
(a gift from Dr. G. M. Rubin)
UAS-Ras
V12
/TM3
(a gift from Dr. N. Perrimon)
vvl
utH599
(a gift from Dr. A. Salzberg)
wg
CX4
(BDSC 2980)
Antibodies used for immunohistochemistry are;
Rabbit anti-GFP (invitrogen)
Rabbit anti-LacZ (Capel)
Rabbit anti-Trh (this study)
Double fluorescent labeling with RNA probes and antibodies was carried out as described (
Goto and Hayashi 1997
).
DNA clones used for in situ RNA detection are;
caup
(Drosophila Genomics Resource Center, DGRC)
trh
(Drosophila Genomics Resource Center, DGRC)
vvl
(a gift from Dr. J. Casanova)
Confocal images were taken by Bio-Rad (Hercules, CA) MRC1024, Olympus (Japan) Fluoview 1000 or Zeiss (Germany) LSM800. Images were processed by ImageJ and figures were prepared with Adobe Photoshop and Illustrator.
Data availability
All the data are available.
Acknowledgements
We thank the members of the fly community who isolated, characterized or distributed fly strains, antibodies or DNA clones. Especially, we thank Drs. C. A. Berg, S. Campuzano, J. Casanova, S. Hayashi, W. Janning, A. S. Manoukian, K. Moses, J. Nambu, A. Nose, N. Perrimon, G.M. Rubin, S. Russel, A. Salzberg, DGRC and BDSC for directly sharing fly strains and DNA clones. We thank Flybase for the Drosophila genomic resources. We thank the Stockholm University Imaging Facility and MBW fly services. We thank V. Tsarouhas for microscope help. Special thanks to Y. Emori and F. Ui-Tei for help in maintaining fly strains after the retirement of K. Saigo.
This work was funded by the Ministry of Education, Culture, Sport, Science and Technology of Japan to K.S. and the Swedish Research Council, the Swedish Cancer Society and German Research Foundation to C.S.
Additional information
Author contribution
RM, conceived the project, designed the experiments, performed experiments, interpreted data, drafted the manuscript with inputs from CH, writing, figure preparation
CH, performed experiments, figure preparation, writing
KS, funding acquisition, provided experiment and analysis tools
CS, funding acquisition, provided experiment and analysis tools, writing
Funding
Cancerfonden (Swedish Cancer Society)
Christos Samkovlis
Vetenskapsrådet (VR)
Christos Samkovlis
Deutsche Forschungsgemeinschaft (DFG)
Christos Samkovlis
Ministry of Education, Culture, Sports, Science and Technology (MEXT)
Kaoru Saigo
References
Fgfr2 is required for limb outgrowth and lung-branching morphogenesis
Proceedings of the National Academy of Sciences of the United States of America
96
:11895–11899
PubMed
Google Scholar
Genetic control of epithelial tube size in the Drosophila tracheal system
Development
127
:3271–3282
PubMed
Google Scholar
. rhomboid, a gene required for dorsoventral axis establishment and peripheral nervous system development in Drosophila melanogaster
Genes & development
4
:190–203
PubMed
Google Scholar
Cross-regulatory interactions among tracheal genes support a co-operative model for the induction of tracheal fates in the Drosophila embryo
Mechanisms of development
91
:271–278
https://doi.org/10.1016/s0925-4773(99)00315-9
PubMed
Google Scholar
The RhoGAP crossveinless-c links trachealess and EGFR signaling to cell shape remodeling in Drosophila tracheal invagination
Genes & development
20
:1817–1828
PubMed
Google Scholar
Identification of the first invertebrate interleukin JAK/STAT receptor, the Drosophila gene domeless
Current biology : CB
11
:1700–1705
https://doi.org/10.1016/s0960-9822(01)00524-3
PubMed
Google Scholar
Alveolar cell fate selection and lifelong maintenance of AT2 cells by FGF signaling
Nat Commun
13
:7137
PubMed
Google Scholar
The transcription factors KNIRPS and KNIRPS RELATED control cell migration and branch morphogenesis during Drosophila tracheal development
Development
125
:4959–4968
PubMed
Google Scholar
The autosomal FLP-DFS technique for generating germline mosaics in Drosophila melanogaster
Genetics
144
:1673–1679
PubMed
Google Scholar
Trachealess (Trh) regulates all tracheal genes during Drosophila embryogenesis
Developmental biology
360
:160–172
PubMed
Google Scholar
Ventral veinless, the gene encoding the Cf1a transcription factor, links positional information and cell differentiation during embryonic and imaginal development in Drosophila melanogaster
Development
121
:3405–3416
PubMed
Google Scholar
Signalling by the sevenless protein tyrosine kinase is mimicked by Ras1 activation
Nature
355
:559–561
PubMed
Google Scholar
In situ activation pattern of Drosophila EGF receptor pathway during development
Science
277
:1103–1106
PubMed
Google Scholar
MAP kinase in situ activation atlas during Drosophila embryogenesis
Development
124
:3535–3541
PubMed
Google Scholar
Branching morphogenesis of the Drosophila tracheal system
Annual review of cell and developmental biology
19
:623–647
PubMed
Google Scholar
The Drosophila FGF-R homolog is expressed in the embryonic tracheal system and appears to be required for directed tracheal cell extension
Genes & development
5
:697–705
PubMed
Google Scholar
Specification of the embryonic limb primordium by graded activity of Decapentaplegic
Development
124
:125–132
PubMed
Google Scholar
The pruned gene encodes the Drosophila serum response factor and regulates cytoplasmic outgrowth during terminal branching of the tracheal system
Development
122
:1353–1362
PubMed
Google Scholar
Drosophila unpaired encodes a secreted protein that activates the JAK signaling pathway
Genes & development
12
:3252–3263
PubMed
Google Scholar
Lung development: orchestrating the generation and regeneration of a complex organ
Development
141
:502–513
PubMed
Google Scholar
Functional subdivision of trunk visceral mesoderm parasegments in Drosophila is required for gut and trachea development
Development
130
:439–449
PubMed
Google Scholar
The torso receptor tyrosine kinase can activate Raf in a Ras-independent pathway
Cell
81
:63–71
https://doi.org/10.1016/0092-8674(95)90371-2
PubMed
Google Scholar
Study of the posterior spiracles of Drosophila as a model to understand the genetic and cellular mechanisms controlling morphogenesis
Developmental biology
214
:197–210
PubMed
Google Scholar
Tubulogenesis in Drosophila: a requirement for the trachealess gene product
Genes & development
10
:103–117
PubMed
Google Scholar
Regulation of Drosophila tracheal system development by protein kinase B
Developmental cell
1
:817–827
https://doi.org/10.1016/s1534-5807(01)00090-9
PubMed
Google Scholar
FGFR2b signalling restricts lineage-flexible alveolar progenitors during mouse lung development and converges in mature alveolar type 2 cells
Cell Mol Life Sci
79
:609
PubMed
Google Scholar
Mammalian tracheal development and reconstruction: insights from in vivo and in vitro studies
Development
148
PubMed
Google Scholar
. breathless, a Drosophila FGF receptor homolog, is essential for migration of tracheal and specific midline glial cells
Genes & development
6
:1668–1678
PubMed
Google Scholar
Mitotic cell rounding accelerates epithelial invagination
Nature
494
:125–129
PubMed
Google Scholar
Two-step regulation of trachealess ensures tight coupling of cell fate with morphogenesis in the Drosophila trachea
eLife
8
PubMed
Google Scholar
Organogenesis in a dish: modeling development and disease using organoid technologies
Science
345
:1247125
PubMed
Google Scholar
. ventral veinless, a POU domain transcription factor, regulates different transduction pathways required for tracheal branching in Drosophila
Development
124
:3273–3281
PubMed
Google Scholar
EGF signalling regulates cell invagination as well as cell migration during formation of tracheal system in Drosophila
Development genes and evolution
209
:174–179
PubMed
Google Scholar
The Development of Drosophila melanogaster
Plainview, N.Y
:
Cold Spring Harbor Laboratory Press
Google Scholar
A resource for manipulating gene expression and analyzing cis-regulatory modules in the Drosophila CNS
Cell reports
2
:1002–1013
PubMed
Google Scholar
The Intersection of the Extrinsic Hedgehog and WNT/Wingless Signals with the Intrinsic Hox Code Underpins Branching Pattern and Tube Shape Diversity in the Drosophila Airways
PLoS genetics
11
:e1004929
PubMed
Google Scholar
Multipotent versus differentiated cell fate selection in the developing Drosophila airways
eLife
4
PubMed
Google Scholar
Fgf-10 is required for both limb and lung development and exhibits striking functional similarity to Drosophila branchless
Genes & development
12
:3156–3161
PubMed
Google Scholar
Ras-independent activation of ERK signaling via the torso receptor tyrosine kinase is mediated by Rap1
Current biology : CB
15
:366–370
PubMed
Google Scholar
Pri micropeptide functions as a cell-intrinsic timer controlling the transient phase of cell fate induction
Proceedings of the National Academy of Sciences of the United States of America
123
:e2511138123
PubMed
Google Scholar
Identification of MAPK phosphorylation sites and their role in the localization and activity of hypoxia-inducible factor-1alpha
The Journal of biological chemistry
281
:33095–33106
PubMed
Google Scholar
A wave of EGFR signaling determines cell alignment and intercalation in the Drosophila tracheal placode
Development
134
:4273–4282
PubMed
Google Scholar
A Drosophila homolog of cadherin associated with armadillo and essential for embryonic cell-cell adhesion
Developmental biology
165
:716–726
PubMed
Google Scholar
A Switch-like Activation Relay of EGFR-ERK Signaling Regulates a Wave of Cellular Contractility for Epithelial Invagination
Developmental cell
46
:162–172
PubMed
Google Scholar
Transcriptional regulation of breathless FGF receptor gene by binding of TRACHEALESS/dARNT heterodimers to three central midline elements in Drosophila developing trachea
Development
124
:3975–3986
PubMed
Google Scholar
Input from Ras is required for maximal PI(3)K signalling in Drosophila
Nature cell biology
8
:1298–1302
PubMed
Google Scholar
Generating lineage-specific markers to study Drosophila development
Developmental genetics
12
:238–252
PubMed
Google Scholar
Tools for neuroanatomy and neurogenetics in Drosophila
Proceedings of the National Academy of Sciences of the United States of America
105
:9715–9720
PubMed
Google Scholar
Vascular heterogeneity and specialization in development and disease
Nat Rev Mol Cell Biol
18
:477–494
PubMed
Google Scholar
The dynamics of tubulogenesis in development and disease
Development
152
PubMed
Google Scholar
Development of the Drosophila tracheal system occurs by a series of morphologically distinct but genetically coupled branching events
Development
122
:1395–1407
PubMed
Google Scholar
Genetic control of epithelial tube fusion during Drosophila tracheal development
Development
122
:3531–3536
PubMed
Google Scholar
Secreted Spitz triggers the DER signaling pathway and is a limiting component in embryonic ventral ectoderm determination
Genes & development
9
:1518–1529
PubMed
Google Scholar
Fgf10 is essential for limb and lung formation
Nature genetics
21
:138–141
PubMed
Google Scholar
Drosophila immune cells transport oxygen through PPO2 protein phase transition
Nature
631
:350–359
PubMed
Google Scholar
An efficient approach to isolate STAT regulated enhancers uncovers STAT92E fundamental role in Drosophila tracheal development
Developmental biology
340
:571–582
PubMed
Google Scholar
Postnatal FGFR-signaling establishes gradients of secretory cell identities along the proximal-distal axis of the lung airways
bioRxiv
:2023.2012.2011.571142
Google Scholar
. branchless encodes a Drosophila FGF homolog that controls tracheal cell migration and the pattern of branching
Cell
87
:1091–1101
https://doi.org/10.1016/s0092-8674(00)81803-6
PubMed
Google Scholar
Convergence of dorsal, dpp, and egfr signaling pathways subdivides the drosophila neuroectoderm into three dorsal-ventral columns
Developmental biology
224
:362–372
PubMed
Google Scholar
Interactions between the EGF receptor and DPP pathways establish distinct cell fates in the tracheal placodes
Development
124
:4707–4716
PubMed
Google Scholar
FGFR-3 and FGFR-4 function cooperatively to direct alveogenesis in the murine lung
Development
125
:3615–3623
PubMed
Google Scholar
An activity gradient of decapentaplegic is necessary for the specification of dorsal pattern elements in the Drosophila embryo
Development
117
:807–822
PubMed
Google Scholar
Genetic control of programmed cell death in Drosophila
Science
264
:677–683
PubMed
Google Scholar
. trachealess encodes a bHLH-PAS protein that is an inducer of tracheal cell fates in Drosophila
Genes & development
10
:93–102
PubMed
Google Scholar
Interaction between Drosophila EGF receptor and vnd determines three dorsoventral domains of the neuroectoderm
Development
125
:3625–3633
PubMed
Google Scholar
The Sox-domain containing gene Dichaete/fish-hook acts in concert with vnd and ind to regulate cell fate in the Drosophila neuroectoderm
Development
129
:1165–1174
PubMed
Google Scholar
Article and author information
Author information
Cite all versions
You can cite all versions using the DOI
10.7554/eLife.111290
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2026,
Matsuda et al.
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
views
0
downloads
0
citations
0
Views, downloads and citations are aggregated across all versions of this paper published by eLife.





