 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库Single Domain Antibody Inhibitors Target the Coiled Coil Arms of theSMC complex
Abstract
Synthetic nanobodies—also called sybodies—have proven valuable for stabilizing conformations of purified proteins, advancing structural and functional studies for example of transmembrane protein complexes. However, their utility in modulating protein function in living cells has remained less well explored. Structural Maintenance of Chromosomes (SMC) complexes facilitate chromosome organization by DNA loop extrusion, a fundamental process in all domains of life. In this study, we target the bacterial SMC complex, Smc-ScpAB, in
Bacillus subtilis
with synthetic nanobodies, aiming to identify key functional regions of the protein complex in a largely unbiased manner. We first isolate sybodies that specifically bind purified Smc-ScpAB and then express them in
B. subtilis
to select binders capable of disrupting Smc-ScpAB function, leading to chromosome segregation defects and cell death. Mapping and biochemical characterization show that the fourteen disruptive sybodies belong to one of three library designs, target the Smc subunit near the same coiled coil arm interface and modulate its ATPase activity in two principal ways, highlighting the mid-region of the Smc coiled coil as critical feature of the DNA folding process. These findings underscore the potential of sybodies—and, by extension, designed binders—as versatile tools for probing dynamic protein function in living cells, with potentially broad applications in cell and synthetic biology.
Introduction
Rapid and specific interference with protein activity and dynamics in living cells is essential for studying and understanding biological mechanisms. However, current approaches—primarily based on small-molecule inhibitors—remain laborious, time-consuming, and costly. The advent of conformation-specific synthetic nanobody (sybody) selection
1
, and more recently, computational design of protein binders
2
, offer unique opportunities for protein-based inhibition and modulation of cellular targets. Here, we generate sybodies against the SMC complex in
B. subtilis
to efficiently inhibit its function within cells.
Structural Maintenance of Chromosomes (SMC) complexes are multi-subunit, ring-shaped ATP-hydrolyzing DNA motors that structure chromosomal DNA by loop extrusion. They are essential for chromosome organization and segregation, gene expression, DNA repair, and defense against non-self DNA across all domains of life
3
. Each SMC protein (Smc in
B. subtilis, bsu
Smc) is a long polypeptide with its N- and C-termini folding together to form a globular ATP binding cassette (ABC) “head” domain. The head is connected to a “hinge” dimerization domain via a ∼50 nm antiparallel coiled coil “arm”, creating an elongated dimer (
Figure 1Aii
and 1Aiii). ATP binding promotes head engagement, bringing the heads together to form a functional ATPase. Non-SMC subunits, a kleisin (ScpA in
B. subtilis
) and a dimer of KITE (ScpB in
B. subtilis
) or two HAWK proteins, bridge the heads, together forming a ring around DNA (
Figure 1Aiii
). In
B. subtilis
, Smc-ScpAB is recruited to the origin of replication (
oriC
) region on the chromosome by ParB, a DNA-binding protein that recognizes centromere-like
parS
sequences (
Figure 2Ai
). Once loaded, Smc-ScpAB translocates at a rate of ∼1 kb/s onto flanking DNA, aligning the two chromosome arms and individualizing nascent sister chromosomes
4
–
6
, likely bypassing obstacles on the chromosome through the SMC hinge gate
7
. Null mutants of
smc
,
scpA
, or
scpB
fail to segregate chromosomes properly and lose viability under conditions promoting rapid growth
5
,
6
,
8
.
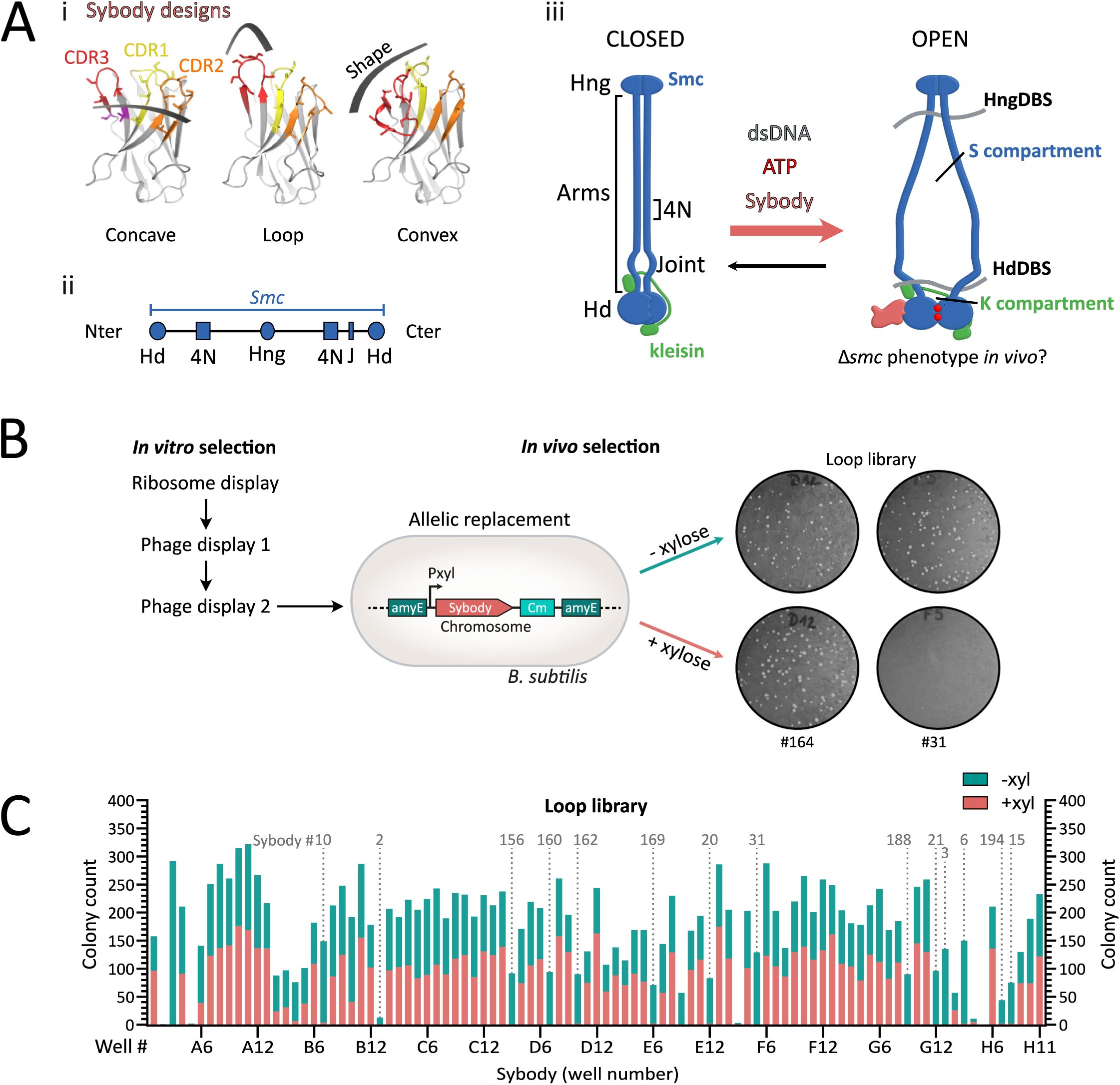
Selection of sybodies targeting
S
mc-ScpAB in
B. subtilis
.
(A) (i)
Structural design and randomization scheme of the three synthetic sybody libraries: concave, loop, and convex. Complementarity-determining regions (CDRs) 1, 2, and 3 are shown in yellow, orange, and red, respectively; randomized residues are shown as sticks. Adapted from
1
.
(ii)
SMC domain organization. The N- and C-terminal regions encode half an ATPase head, while the middle domain constitutes the hinge dimerization domain. The elongated sequence in-between encodes for long helical sequences that will form an antiparallel intramolecular coiled coil. Hd: head, Hng: hinge, 4N: 4N arm-to-arm contact, J: joint.
(iii)
SMC complexes harbor juxtaposed arms in their resting state. Upon ATP binding at the heads, the heads dimerize (head engagement), leading to a conformational change of the entire complex in which the arms open. This state is thought to fit a loop of DNA (not shown) in the S compartment formed by the SMC arms, that will be pushed towards the K compartment formed by the heads and kleisin, after ADP release. We hypothesized that SMC-blocking sybodies can stabilize distinct conformation of the
bsuS
mc-ScpAB complex promoted by ATP hydrolysis and DNA binding, potentially inducing a
Δsmc
-like phenotype
in vivo
and enabling biochemical characterization of trapped conformations
in vitro
. Blue: SMC dimer, Green: kleisin ScpA. The ScpB KITE subunits are not represented for the sake of simplicity. HngDBS: Hinge DNA-binding site, HdDBS: Head DNA-binding site.
(B)
Framework for sybody selection.
In vitro
selection starts with ∼10¹² sybody variants per library subjected to ribosome display for pre-enrichment, followed by two rounds of phage display. For
in vivo
selection, 95 randomly selected sybody genes were integrated into the
B. subtilis
chromosome under the control of a xylose-inducible promoter by allelic replacement at the
amyE
locus. Growth defects on rich medium were tested on ONA agar plates with or without 0.5% xylose. Example shown: Sb164 (loop library) did not affect growth, whereas Nb31 impaired growth upon induction, suggesting interference with
bsuS
mc function.
(C)
Growth assay results for the 95
B. subtilis
strains expressing individual xylose-inducible sybodies of the loop library. Bars show colony counts “without” (green) on top of “with” (pink) xylose. Strains are ordered by their original position in the 96-well plate. Fourteen sybodies consistently impaired colony formation under inducing conditions (marked by dotted lines). Sybody numbers indicated above the plots correspond to selected candidates used in subsequent experiments, numbering according to order of first use. Notably, the strain corresponding to the E09 sybody (Sb018), showed an absence of colonies upon sybody induction. However, this sybody candidate gave intermediate phenotypes in later experiments, which is why it was excluded from detailed analysis.
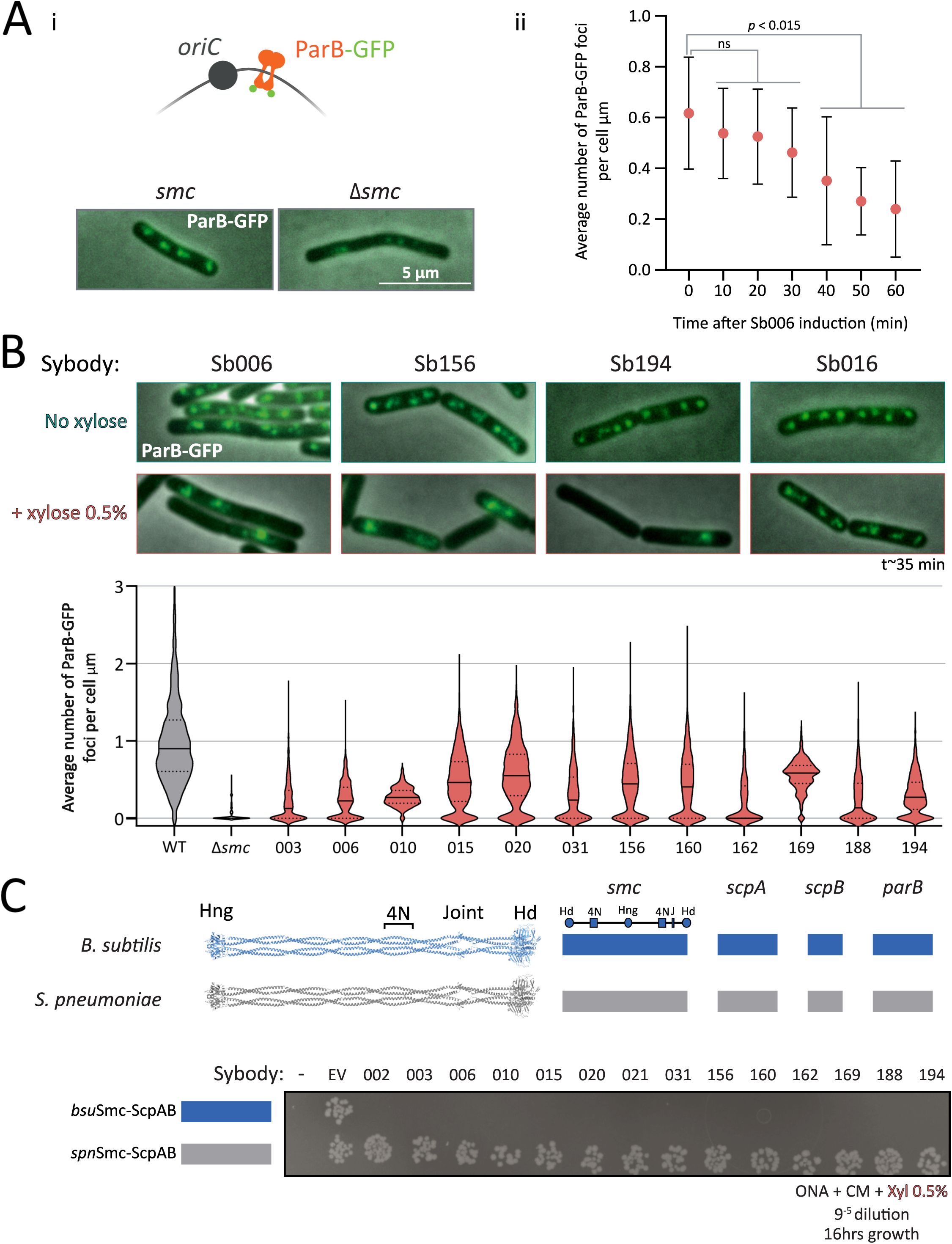
Sybody-induced chromosome segregation defects visualized by ParB-GFP imaging.
(A)
i,
Top
: Schematic illustrating ParB-GFP binding near
oriC
, enabling visualization of origin positioning.
Bottom
: Representative images showing ParB-GFP foci in wild-type and
Δsmc B. subtilis
.
Smc
WT cells typically display 2–4 foci per cell, whereas
Δsmc
cells exhibit reduced foci numbers. ii, Number of ParB-GFP foci per µm of cell length in a strain carrying inducible sybody Sb006. A significant decrease in foci is detected from 40 minutes post-induction (
pₜ₀₋₁₀
= 0.9674;
pₜ₀₋₂₀
= 0.9033;
pₜ₀₋₃₀
= 0.3744; p
ₜ₀₋₄₀
= 0.0145; p
ₜ₀₋₅₀
= 0.0004; p
ₜ₀₋₆₀
< 0.001). Based on this, a standardized induction time of ∼35 minutes was used in subsequent experiments.
(B)
ParB-GFP foci density (foci/µm) in WT,
Δsmc,
and sybody-expressing strains after 35 minutes of xylose induction. Violin plots show distribution per condition; solid lines denote the mean, and dotted lines indicate quartiles. Several hundreds of cells were analyzed (between 351 and 1735), except for
Δsmc
and Sb010 were fewer cells were available (134 and 73, respectively).
(C)
Spot assay to assess colony formation of
B. subtilis
strains
bsu
or
spn
variants of
S
mc-ScpAB and ParB.
Top
: Schematic shows gene origins (blue:
B. subtilis
; grey:
S. pneumoniae
). The leftmost column corresponds to the parental
B. subtilis
strain 1A700 carrying the
S. pneumoniae
genes without chloramphenicol resistance (non-growing). The next spots represent the same strains carrying no sybody gene but the Cm resistance (EV for empty vector). Remaining spots are sybody-expressing strains; sybody numbers are indicated above. Cells were grown for 16 h at 37 °C on ONA supplemented with 0.5% xylose and chloramphenicol. Hd: head, Hng: hinge, 4N: 4N arm-to-arm contact, J: joint.
Several models have been proposed for the mechanism of DNA loop extrusion by SMC complexes, including the “segment capture” model, in which DNA segments are transiently trapped between the SMC arms in partially open states and fused into larger loops through iterative ATP-driven cycles (
Figure 1Aiii
and S1B)
9
–
11
. However, detailed structural understanding and experimental testing
in vivo
are lacking.
Antibodies can block biochemical reactions by stabilizing reaction intermediates. Synthetic single-domain antibodies called sybodies are small, robust antigen-binding proteins that were engineered based on three camelid nanobody structures
1
. They are selected from synthetic libraries that encode a wide diversity of binding surfaces and epitope shapes based on the length and geometry of their CDR3 loop, allowing strong binding of diverse antigen surfaces (
Figure 1Ai
). Their small size (∼15 kDa), stability, and ability to bind transient epitopes make them ideal tools for targeting specific states of biomolecules
12
. Selection is carried out using purified and immobilized Smc-ScpAB by ribosome display and phage display, typically followed by an ELISA-based screening
1
.
Here, we demonstrate that sybodies can be used to interfere with the function of
B. subtilis
Smc-ScpAB
in vivo
. We first isolate binders that specifically target purified Smc-ScpAB
in vitro
and then select those that eliminate Smc function when expressed in
B. subtilis
. Fourteen sybodies were found to disrupt chromosome segregation, mimicking
smc
deletion phenotypes. Biochemical assays confirmed that selected sybodies alter the ATP hydrolysis rate of Smc-ScpAB and its stimulation by DNA addition, likely stabilizing intermediates of the conformational cycle. Mapping experiments using chimeric Smc constructs revealed that most binders, unexpectedly, target the coiled coil, specifically in a defined region near the Smc 4N arm-to-arm contact
13
, highlighting the potential of sybodies and designed binders, as genetic tools to identify essential functional domains and underscoring the importance of coiled coil dynamics for Smc function.
Results
Generation of
bsuS
mc-ScpAB-specific sybodies
To isolate sybodies that impede the function of
B. subtilis
Smc-ScpAB complex, we first performed
in vitro
selection with purified Smc-ScpAB in the presence of 40 bp duplex DNA, and an ATPase-deficient mutant (E1118Q) of
bsuS
mc. These conditions favour ATP-engaged complexes alongside the typically predominant ATP-disengaged rod-shaped state
13
.
bsuS
mc was also biotinylated at the hinge at residue R643C for immobilization (
Figure 1Aii-iii
and S1A)
1
,
14
–
16
. Starting from three synthetic sybody libraries encoding distinct epitope-binding geometries (denoted as concave, loop, convex, respectively,
Figure 1Ai
), we performed one round of ribosome display starting from a large sybody library followed by two rounds of phage display, thereby enriching sybodies that bind to Smc-ScpAB (
Table 1
). From each library, 95 sybody-expressing
E. coli
clones were randomly chosen for
in vivo
characterization. ELISA data revealed that nearly all clones bind purified Smc-ScpAB (
Table 1
). However, the ELISA signals of only few Sybodies showed clear dependence on the presence or absence of ATP and DNA (Table S1).
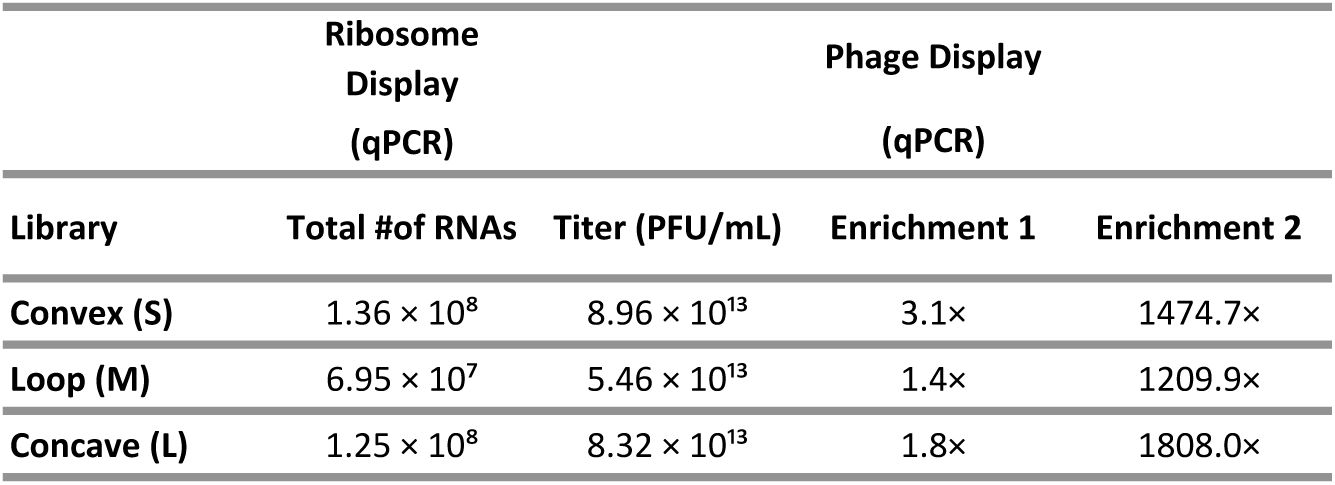
Sybody enrichments at different steps of the selection procedure.
Ribosome display output was quantified by qPCR, while phage display results include final phage titers and enrichment values from rounds one and two, also measured by qPCR.
Identification of
bsuS
mc-ScpAB-blocking sybodies through phenotypic screening in
B. subtilis
We therefore decided to screen for inhibiting sybodies directly in
B. subtilis
cells. Despite containing a conserved disulfide bond, sybodies have been successfully expressed inside cells and were shown to bind their targets in the reducing environment of the cytoplasm, likely owing to their robust folding
17
. To assess whether the selected sybodies interfere with Smc function
in vivo
, each of the 285 sybody sequences was cloned under a xylose-inducible promoter (P
xyl
) into an
E. coli–B. subtilis
shuttle vector and integrated at the
amyE
locus of the wild-type 1A700 strain (
Figure 1B
)
18
. Fourteen sybodies from the loop library failed to yield transformants in the presence (but not absence) of xylose, indicating cell lethality due to sybody inhibition of Smc-ScpAB (
Figure 1C
, S2A-B). These Smc-disruptive sybodies (denoted as Sb002, 003, 006, 010, 015, 020, 021, 031, 156, 160, 162, 169, 188, and 194, respectively) harbor fourteen distinct sequences(Table S2). Intriguingly, no such disruptive sybodies were isolated from the concave and convex libraries (
Figure S2B
), suggesting that the CDR3 geometry as present in the loop library is particularly effective at inhibiting
bsuS
mc-ScpAB. Likely, sybodies of the other libraries bound to other epitopes of
bsuS
mc-ScpAB that are not sensitive to conformational trapping, and it is further possible that they were not stably expressed in
B. subtilis
. However, at least for sybodies from the loop library, we did not notice any obvious correlation between expression levels and phenotypes. Selected sybodies showed similar expression and were appreciably expressed even without inducer (
Figure S4
,
S5
). Notably, tendencies of preferential isolation of binders from one of the three libraries have previously been observed, although at milder levels
1
,
17
,
19
.
Selected sybodies target
bsuS
mc-ScpAB function
in vivo
To determine whether the fourteen sybodies indeed impair chromosome segregation, we used fluorescence microscopy to monitor
oriC
positioning in a
B. subtilis
strain expressing a ParB-GFP fusion protein together with a P
xyl
-inducible sybody gene. In these strains, ParB-GFP marks the replication origin region by binding to
parS
sites near
oriC
(
Figure 2Ai
). As expected, the
Δsmc
strain displayed fewer ParB-GFP foci and elongated cells (
Figure 2Ai
) (Table S3). These elongated cells are known to harbour expanded nucleoids, consistent with delayed
oriC
separation rather than delayed DNA replication
6
,
20
. A time course experiment using sybody Sb006 revealed chromosome organization defects as early as 30 minutes post induction, with a significant reduction in ParB-GFP foci per µm cell length at 40 minutes (
p
= 0.0145) (
Figure 2Aii
). Subsequent imaging was performed ∼35 minutes after induction of sybody expression.
All disruptive sybodies reduced ParB-GFP foci density compared to the control (1.03 foci/µm), with values ranging from 0.56 foci/µm (Sb020) to 0.22 foci/µm (Sb003). These defects were all robustly detected, while being less severe than the Δ
smc
mutant (∼0.05 foci/µm), conceivably due to the short induction time or incomplete inhibition. Milder defects were also observed without induction, likely attributed to leaky expression from the P
xyl
promoter (
Figure S3A
). Sybody expression also caused cell elongation and growth delays, both hallmarks of impaired Smc activity, consistent with chromosome segregation defects that delay cell division (
Figure S3B–C
). Altogether, these results show that all selected sybodies induce
Δsmc
-like phenotypes likely by interfering with
bsu
Smc-ScpAB activity
in vivo
.
Selected sybodies specifically target
B. subtilis
Smc-ScpAB
To test whether sybodies specifically target
B. subtilis
Smc-ScpAB, we assessed their effects in a strain expressing
Streptococcus pneumoniae
(
Spn
) Smc-ScpAB and ParB proteins in place of the endogenous
B. subtilis
proteins. The
Spn
sequences can functionally replace the corresponding
Bsu
sequences despite significant sequence divergence (∼38% sequence identity)
21
. Strikingly, none of the sybodies impaired growth in this background, even though a cognate ParAB pair is absent in this strain (
S. pneumoniae
lacks
parA
), sensitizing cells to chromosome segregation defects (
Figure 2C
). These results confirm that the sybodies are specific to
bsu
Smc-ScpAB and that off-target toxicity is not noticeable.
bsuS
mc-ScpAB-disrupting sybodies target two distinct coiled coil regions adjacent to the
bsuS
mc 4N arm-to-arm contact
To map the sybody-binding site on the
bsuS
mc-ScpAB complex, we utilized five chimeric Smc constructs (Smc Chimera 1-5), in which the hinge and progressively longer segments of the adjacent coiled coils were replaced with the corresponding sequences from the
S. pneumoniae
Smc protein (
Figure 3A
). Chimeric junctions were designed to preserve coiled coil integrity based on available crystal structures, coiled coil predictions by DeepCoil, and AlphaFold2 structural models
1
,
22
,
23
. All chimeric Smc strains retained the native
B. subtilis scpA
,
scpB
, and
parB
genes and were viable under conditions promoting fast growth, demonstrating proper protein folding and functioning of chimeric Smc-ScpAB complexes (
Figure S6A
).
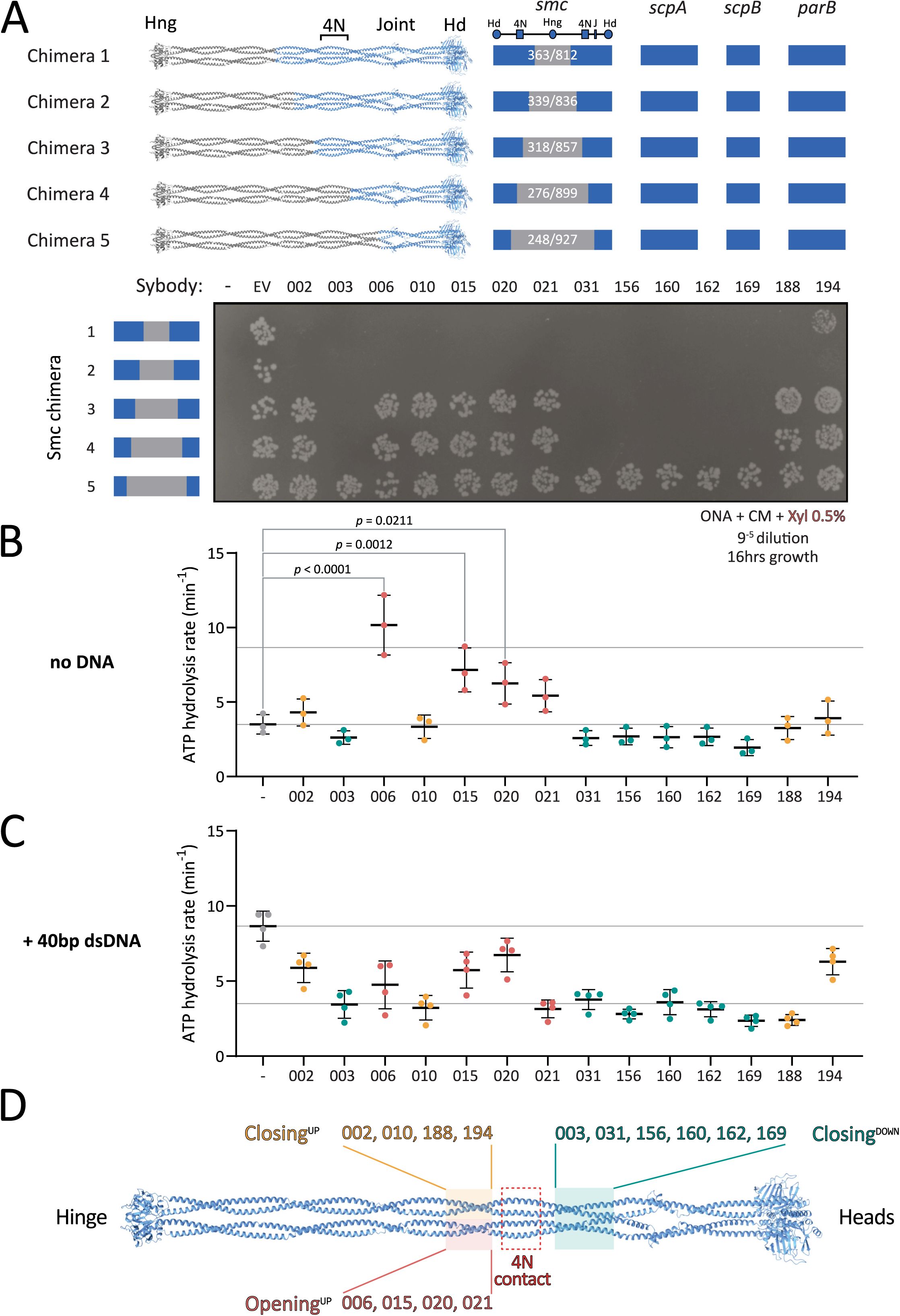
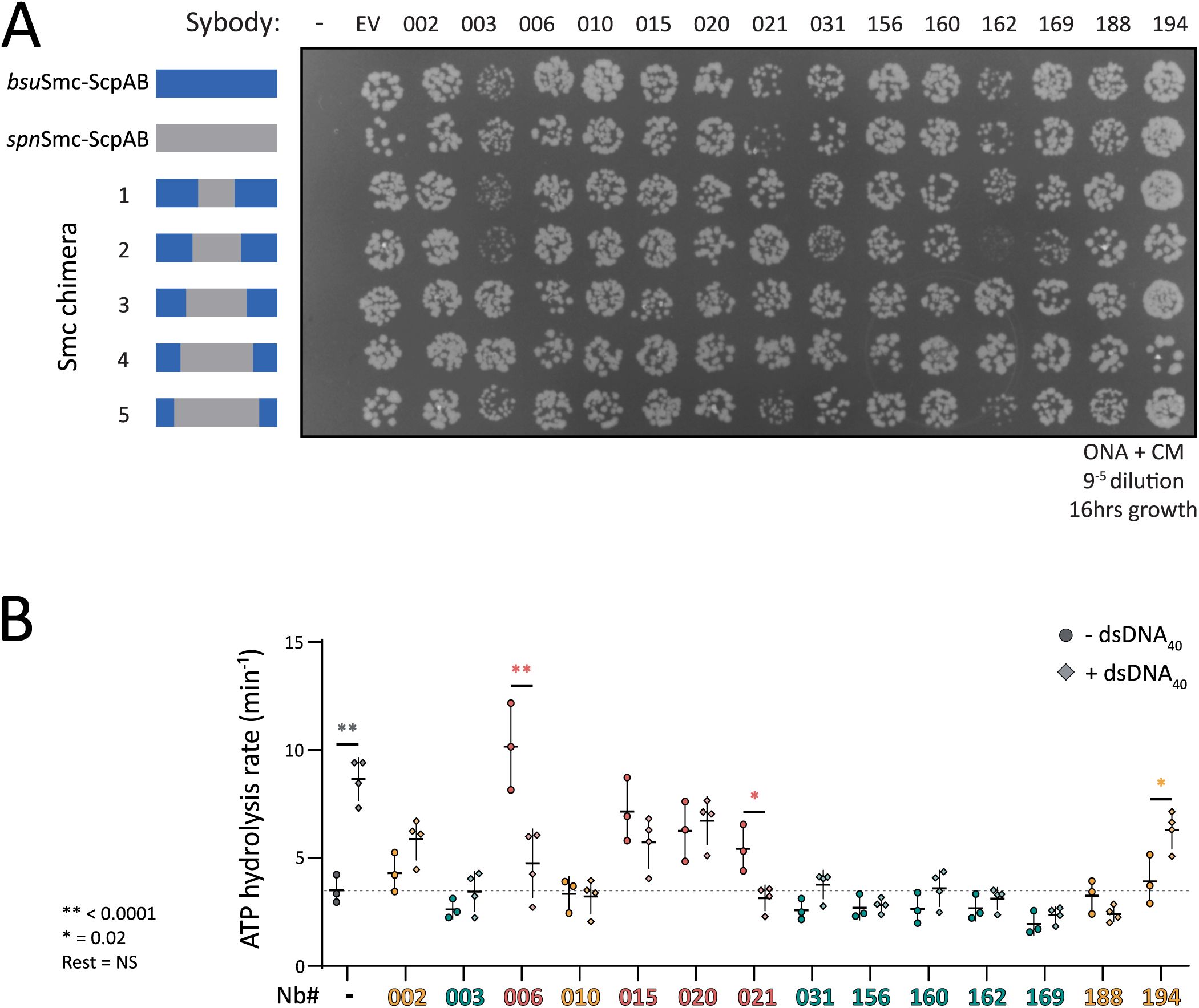
Mapping sybody binding sites on Smc-ScpAB.
(A)
Colony formation of
B. subtilis
strains expressing chimeric Smc-ScpAB complexes comprising
S. pneumoniae
and
B. subtilis
sequences. The schematic above the spot assay depicts the species origin of the
smc
gene, the
scpAB
operon and
parB
gene (blue:
B. subtilis
; grey:
S. pneumoniae
). “–” indicates no insertion at the
amyE
locus; “EV” refers to the empty vector control containing only the chloramphenicol resistance cassette that was inserted in
amyE
; numbered labels correspond to sybodies. Cells were spotted on rich medium (ONA) supplemented with chloramphenicol and xylose and incubated for 16 h at 37 °C. Hd: head, Hng: hinge, 4N: 4N arm-to-arm contact, J: joint.
(B)
ATP hydrolysis rates of
bsu
Smc-ScpAB in the presence of sybodies but absence of DNA. Significant effects by one-way ANOVA are indicated by p values.
(C)
ATPase rates in the presence of 40 bp dsDNA. All sybodies significantly reduced DNA-stimulated ATP hydrolysis. Reported p-values: Sb020 (p = 0.340), Sb194 (p = 0.0049), Sb002 (p = 0.0007), Sb015 (p = 0.0003); all others, p < 0.0001.
(D)
Schematic summary of sybody binding sites mapped onto the Smc dimer, categorized by their effect on ATPase activity. Sybodies are grouped based on functional impact and mapped to corresponding structural regions: pink/yellow boxes indicate residues 318–339 and 836–857; green boxes mark residues 248–276 and 899–927; and the red box highlights the 4N contact region (approx. residues 290–320).
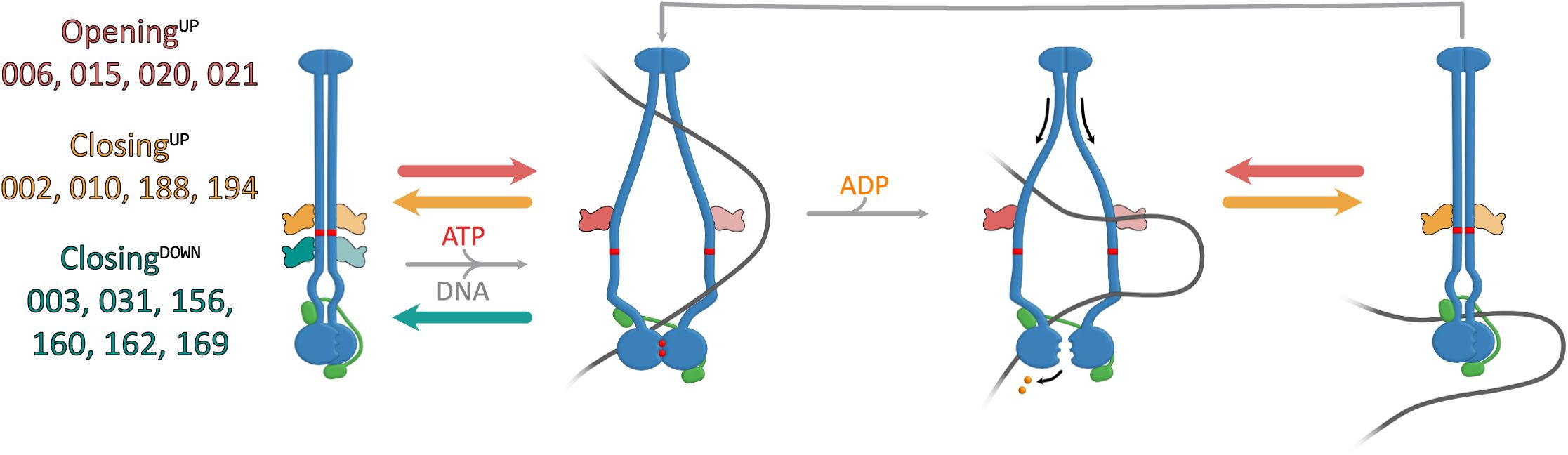
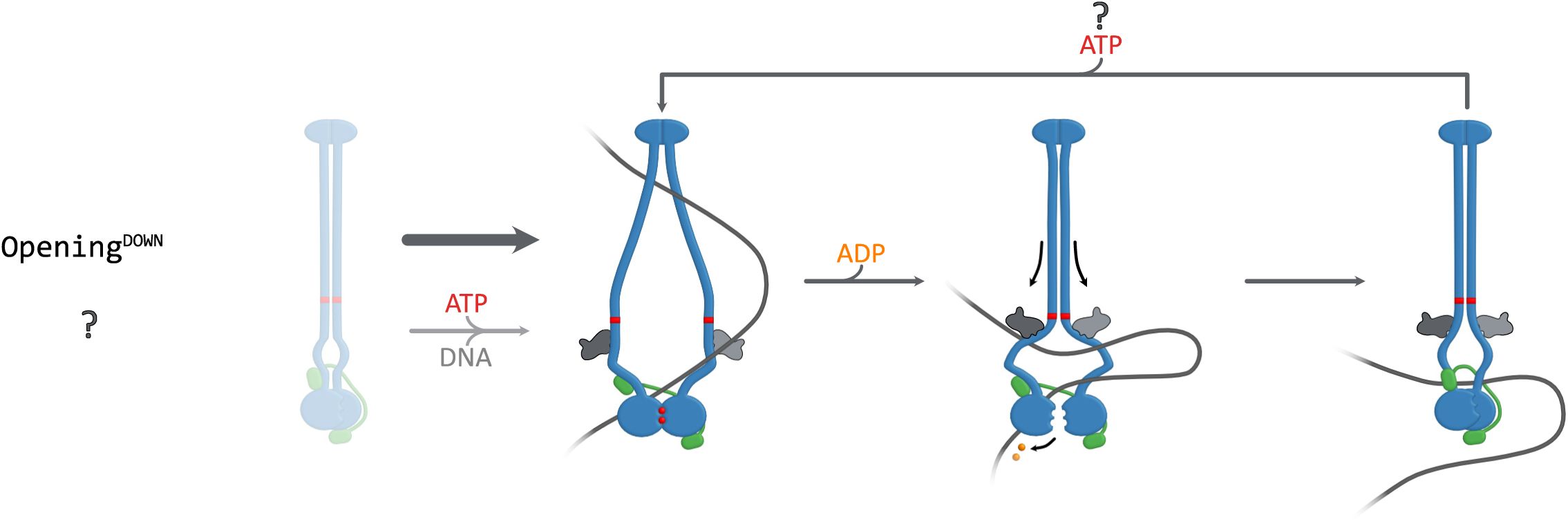
Proposed models for sybody interactions with
bsuS
mc-ScpAB during the ATPase cycle.
(A)
Model for sybodies of the Opening
UP
, Closing
UP
and Closing
DOWN
group. Opening
UP
sybodies, likely prevent complete arm closure. In the presence of DNA, these sybodies may stabilize a suboptimal conformation for head disengagement, resulting in reduced ATPase activity (not shown here). Closing
UP
and Closing
DOWN
sybodies stabilize a closed-arm conformation. These likely hinder DNA segments from entering the inter-arm space and accessing the hinge-proximal DNA binding site.
Chimeras 1 and 2, which carry the central hinge and coiled coil sequences of
Spn
origin (residues 363-812 and 339-836, respectively), showed the same sybody-induced growth defects as the wild-type strain, indicating that the sybodies bind further away from the hinge, within the remaining
B. subtilis
portion comprising of the Smc head and the first ∼230 coiled coil residues, and ScpAB (
Figure 3A
). By contrast, Chimera 5 (residues 248–927 of
Spn
origin), supported robust growth even in the presence of the
bsu
Smc-disrupting sybodies, indicating that the region comprising the heads and first ∼50 residues of the coiled coil (including the joint) does not comprise the binding site. The binding region for the sybodies is thus located on the coiled coil between residues 248 and 339 (or 836 and 927), demonstrating that all fourteen sybodies bind to a central region of the coiled coil.
Chimera 3 (
spn
Smc residues 318-857) and chimera 4 (
spn
Smc residues 276-899) revealed two distinct sybody responses. Sb003, 031, 156, 160, 162, and 169 impaired growth of both strains, mapping their binding sites to
B. subtilis
residues 248-276 (and 899-927), present in chimeras 1–4 but absent in chimera 5. In contrast, Sb002, 006, 010, 015, 020, 021, 188, and 194 had no impact on the viability of chimera 3 or 4, suggesting they target residues 318-339/836-857 (
Figure 3A
, 3D).
In sum, these experiments mapped the binding sites of all fourteen disruptive sybodies to one of two ∼twenty amino acid segments in the central region of the
bsuS
mc coiled coil (
Figure 3D
). These regions harbor relatively poorly conserved sequences but flank the 4N arm-arm contact (∼at residue 295), previously proposed to serve as an essential structural switch between closed and open conformations
13
. These results reveal that all isolated sybodies target the same functional element of the Smc arms, and that interfering with this element disrupts Smc-dependent chromosome organization
in vivo
.
Smc-disrupting sybodies affect the ATPase activity in one of two ways
To investigate how sybodies may influence
bsuS
mc-ScpAB, we next measured ATP hydrolysis rates using purified proteins in the presence and absence of DNA. SMC ATPase activity serves as an indirect readout for state transitions, including arm opening and head engagement. In absence of DNA,
bsuS
mc-ScpAB hydrolyzes ATP at ∼4 ATP/min/Smc
13
. DNA binding stimulates ATP hydrolysis to ∼9 ATP/min/Smc, presumably by DNA binding promoting arm opening and head engagement, creating a more open conformation.
All disruptive sybodies measurably affected the ATP hydrolysis rate of
bsuS
mc-ScpAB. Ten maintained a near-basal ATPase rate even in presence of DNA (Sb002, Sb003, Sb010, Sb031, Sb156, Sb160, Sb162, Sb169, Sb188, and Sb194) (
Figure 3B-C
and S6B). This suggests that they stabilize the complex in a closed or partially closed conformation, thus hindering DNA-dependent ATPase stimulation. Notably, in this group, Sb002 and Sb194 showed slightly elevated ATPase rates with DNA, hinting at a mildly more flexible conformation.
In contrast, the other sybodies (Sb006, Sb015, Sb020, and Sb021) stimulated ATP hydrolysis in the absence of DNA, with Sb006 reaching levels comparable to DNA-induced activation (∼10 ATP/min). This implies that these sybodies stabilize Smc-ScpAB in a more open state promoting head engagement. Curiously, addition of DNA in the presence of Sb006 and Sb021 reduced—rather than stimulated—ATPase activity (
Figure 3B-C
), suggesting that concurrent DNA and sybody binding may trap the complex in a non-productive conformation, perhaps by favoring the open state excessively. These findings are consistent with the idea that 4N contact region plays a critical role in arm opening and ATPase stimulation.
Interestingly, the observed ATPase profiles correlate with distinct binding regions along the Smc arms. All ATPase-stimulating sybodies (Sb006, Sb015, Sb020, Sb021) bind above the 4N contact (“Opening
UP
”), while sybodies that hinder DNA stimulation fall into two subgroups: those binding above the 4N contact (Sb002, Sb010, Sb188, Sb194—“Closing
UP
”) and those binding below it (Sb003, Sb031, Sb156, Sb160, Sb162, Sb169—“Closing
DOWN
“) (
Figure 3D
). These results indicate that the sybodies perturb
bsuS
mc-ScpAB in two principal ways: either by favoring the opening of the arms or by constraining the arms in a more closed state. The results highlight arm opening and closure as central regulatory features of bacterial Smc-ScpAB
9
,
11
,
24
,
25
and confirm the key role of the 4N arm contact in balancing the opening and closing of SMC arms
13
.
Discussion
Over the past three decades, nanobodies and their synthetic counterparts have transformed biomedical research and structural biology by enabling the targeting and stabilization of transient protein conformations. Despite these advances, their application to probing protein function in living cells has remained limited
17
,
26
. Here, we expand the utility of sybody selection by employing it as a tool to screen for genetic probes targeting the function of selected molecular machines
in vivo
. This strategy combines the advantages of controlled biochemical reconstitution with the ability to study the function of proteins in their native context. Importantly, sybody selection requires no prior knowledge of vulnerable regions, providing an unbiased means to interrogate protein function that is difficult to achieve through rational design. Our study based on the
B. subtilis
Smc-ScpAB DNA motor as a model uncovers a central regulatory region in the coiled coil arms of Smc and highlight how distinct arm conformations shape SMC function in loop extrusion and chromosome segregation.
Revealing Smc arm dynamics through synthetic binders
All fourteen Smc-disrupting sybodies share the loop library design. Moreover, their Smc binding sites map to the two short coiled coil segments flanking the conserved 4N arm-to-arm contact, a region that has previously been implicated in key conformational transitions during arm opening
13
. While the fourteen sybodies harbour different epitope binding sequences, we suspect that they may all bind to the Smc dimer in an analogous manner. With the long CDR3 loops known to bind narrow structural cavities
12
,
27
, we speculate that these sybodies may intercalate between the two coiled coil arms (
Figure 3D
). Structural studies are needed to test this hypothesis directly, however, our attempts at X-ray crystallography and electron microscopy of Smc-sybody complexes have failed so far.
Notably, while the coiled coil dimer appears similarly structured from the elbow to the hinge
9
(
Figure 3D
), no Smc-disrupting sybodies mapped to regions further away from the 4N contact (
Figure 3D
). This indicates that opening and closure of the Smc arms might be dispensable at these places, although formal testing would require isolating binders efficiently targeting these regions, potentially facilitated by binder design, and expressing them in
B. subtilis
. This parallels with recent findings with the bacterial SMC defense system Wadjet, where hinge-proximal arm opening is dispensable for loop extrusion, but essential for obstacle bypass
7
. Alternatively, these arm-to-arm contacts might be more stable and thus less likely to be interfered with by sybody binding. Similarly, sybodies targeting below the 4N contact all reduce the ATPase rate (or rather its DNA stimulation), indicating that artificial opening at this position may not disrupt Smc function (
Figure S7
). Again, these hypotheses need to be tested more directly. We envision that the design of protein binders may allow us to confirm these observations and further dissect the underlying mechanisms.
This work establishes sybodies as precision tools for blocking ATP-driven machines inside bacterial cells. By targeting allosteric control points in Smc, they enable mechanistic dissection of loop extrusion and open new avenues for studying dynamic complexes
in vivo
. More broadly, the study demonstrates how synthetic binders can trap or block conformations of active chromatin-associated machines, providing a powerful means to probe their mechanisms in living cells. Looking ahead, the rational design of protein binders with tailored geometry and allosteric potential could allow researchers to manipulate and visualize specific conformational states with even greater control and refinement, across both prokaryotic and eukaryotic organisms, while the generation and
in vivo
screening of sybodies remains an attractive approach to gain unexpected biological insights in an unbiased manner.
Material and Methods
Bacillus subtilis
strains construction and growth
B. subtilis
strains used in this study were derived from the parental strain 1A700. Allelic replacement was achieved via double-crossover recombination at the endogenous
smc
locus using the natural competence method described by
18
,
28
. Chromosome-integration of sybody genes were performed by transforming a vector containing an
amyE h
omologous regions on each side of the recombinant gene of interest, as described in
18
. Transformants were selected on ONA solid medium containing the appropriate antibiotics. Following transformation, strains were purified by single colony isolation and verified through a combination of marker testing, phenotype assessment, PCR, and Sanger sequencing of the
smc
locus, when appropriate. For spot assays, cells were cultured to stationary phase in liquid LB medium, and 9
-
2
and 9
-
5
dilutions were then spotted onto solid ONA medium with chloramphenicol (5 ug/mL) and xylose (0.5% w/v) when induction of the P
xyl
promoter was required. The strains, plasmids, and oligonucleotides used in this study are listed in Table S4, Table S5 and Table S6, respectively.
For viability assessment, 200 LB medium was inoculated with a 1:1000 dilution of a dense
B. subtilis
culture and grown to stationary phase. The culture was then serially diluted in LB medium. From each dilution (9⁻¹ to 9⁻⁸), 5 µL was spotted onto LB agar plates containing the appropriate antibiotic selection. Plates were incubated, and viability was assessed by imaging the spots at 16- and 24-hours post-incubation.
Sybody selection
Sybody selection was performed following the protocol by
16
, with modifications to include binders specific for the ATP/DNA-stabilized “open” form of the
bsuS
mc-ScpAB complex.
Ribosome Display
The standard WTB buffer (50 mM Tris/acetate pH7.4, 150 mM NaCl, 50 mM MgAc2) and its derivatives were adjusted to 50 mM NaCl to facilitate
bsuS
mc-ScpAB binding to dsDNA. The target complex was assembled by mixing 500 nM
bsuS
mc(C119S, C437S, C826S, E1118Q, R643C)-ScpAB, 2 mM ATP, and 5 µM 40 bp dsDNA in WTB-D-BSA buffer (WTB including 0.5% (w/v) BSA and 0.1% Tween-20), incubated for 15 min at room temperature (RT). The ribosome display panning solution was supplemented with 2 mM ATP and 5 µM dsDNA to maintain binding conditions. Solution panning and biotinylated target capture were conducted at RT to optimize selection for the “open” complex. Washing steps incorporated 2 mM ATP and 5 µm 40 bp dsDNA, while elution, reverse transcription, and cDNA amplification followed the original protocol. Reverse transcribed RNA molecules were quantified by qPCR.
First round of phage display
Phagemid libraries were cloned and electroporated into
E. coli
SS320 as previously described
16
. Phage production was performed using M13KO7 helper phage. The phage display buffer (TBSM) consisted of 20 mM Tris-HCl (pH 7.4), 50 mM NaCl, and 2 mM MgCl2. Target preparation involved incubating 500 nM biotinylated
bsuS
mc(C119S, C437S, C826S, E1118Q, R643C)ScpAB with 2 mM ATP and 5 µM dsDNA in TBSM-BSA-D (TBSM including 0.5% (w/v) BSA and 0.05% Tween-20). Phages (10¹²/ml) were incubated with the target at 50 nM for 20 min at RT before immobilization on neutravidin-coated plates. Washing (with added 2 mM ATP and 5 µm 40 bp dsDNA) and elution otherwise followed the original protocol, with enrichment assessed by qPCR. Amplified phages were used for a second selection round.
Second round of phage display
Purified phages from the first round were used at 5 × 10¹³ phages/ml, and the target was prepared as in the first round. Phage-target binding occurred at 50 nM in the presence of ATP and dsDNA, followed by capture on magnetic beads. A competition step using non-biotinylated
bsuS
mc-ScpAB (1 µM) was included before washes with TBSM-D. Phages were eluted and enrichment was determined by qPCR. Phagemids were purified, sub-cloned into the pNb_init vector, and transformed into
E. coli
MC1061.
Growth curve analyses
Growth curves were generated from
B. subtilis
strains grown overnight to exponential phase in LB medium supplemented with 0.5% glucose, incubated at 30°C with shaking. The following day, fresh 5 mL LB cultures were inoculated with the overnight culture to an OD of 0.005 and incubated at 37°C with shaking until reaching an optical density (OD) of 0.05. Subsequently, each culture was subjected to a twofold dilution in a 96-well plate (Costar #3596), achieving a final volume of 200 µL per well. Where necessary, cultures were induced with 0.5% xylose. Plates were incubated at 37°C with continuous shaking in a Thermo Scientific Multiskan FC plate reader. Growth curves were determined by light scattering at 620 nm. The BactEXTRACT app was used to perform Analysis and visualization of the data
29
.
Protein purification
Purification of
bsuS
mc and biotinylated
bsuS
mc(C119S, C437S, C826S, E1118Q, R643C)
bsuS
mc proteins were purified according to
14
. pET-22 or pET-28 plasmids encoding the
Smc
recombinant sequences were transformed into
E. coli
BL21-Gold (DE3) cells. Protein expression was carried out in ZYM-5052 autoinduction medium for 23 hours at 24°C. The cells were harvested and resuspended in lysis buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1 mM EDTA, 1 mM DTT, 10% (w/v) sucrose) supplemented with protease inhibitor cocktail. Cell lysis was achieved by sonication, and the lysate was clarified by centrifugation. The supernatant was filtered through a 0.45 µm membrane and loaded onto two HiTrap Blue HP 5 mL columns connected in series. Elution was performed using lysis buffer with 1 M NaCl. The main peak fractions were pooled and diluted with salt-free buffer (50 mM Tris-HCl, pH 7.5; 1 mM EDTA; 1 mM DTT) to a final conductivity equivalent to 50 mM NaCl (∼8 mS/cm). This diluted sample, supplemented with protease inhibitor cocktail, was applied to a HiTrap Heparin HP 5 mL column, and the proteins were eluted using a linear gradient up to 2 M NaCl. The main peak fractions (∼ 5 mL) were collected and subjected to further purification by gel filtration using an XK 16/70 Superose 6 PG column equilibrated with 50 mM Tris-HCl, pH 7.5, 200 mM NaCl, 1 mM EDTA, and 1 mM TCEP. The peak fractions were collected, concentrated using a Vivaspin 15 10K MWCO filter, flash frozen in liquid nitrogen, and stored at −80°C. Protein concentration was determined by absorbance, utilizing theoretical molar absorption and molecular weight values.
For the biotinylated
bsu
Smc(C119S, C437S, C826S, E1118Q, R643C) protein used in the sybody selection, the protein was first purified as described above, except that the reducing agent (TCEP) was omitted from the final gel filtration buffer. Labelling was performed by incubating 600 µL of
bsuS
mc(C119S, C437S, C826S, E1118Q, R643C) (83 µM in 50 mM Tris-HCl pH 7.5, 200 mM NaCl) with 1 mM PEG2-biotin maleimide for 10 minutes at 4 °C. The reaction was quenched by addition of 0.5 mM 2-mercaptoethanol. Excess label was removed using Zeba spin desalting columns (Thermo Fisher) in multiple parallel runs due to volume limitations. The final yield was 600 µL at 72.35 µM (subunit concentration). Labelled
bsuS
mc(C119S, C437S, C826S, E1118Q, R643C) was loaded onto a Superose 6 10/300 Increase column pre-equilibrated in 50 mM Tris-HCl pH 7.5, 200 mM NaCl. Peak fractions were pooled and mixed with ScpA and ScpB at a 1:1:2 molar ratio relative to the
bsuS
mc dimer. The final concentration of the full complex was adjusted to 5.7 µM. Complexes were aliquoted in 100 µL portions, flash-frozen in liquid nitrogen, and stored at −80 °C.
Purification of
bsu
ScpA
ScpA was purified using the method described by
25
.
E. coli
BL21-Gold (DE3) cells, transformed with a pET-28 derived plasmid encoding the ScpA protein, were used for expression. Cultivation was performed in ZYM-5052 autoinduction medium at 16°C for 28 hours. Cells were then harvested and resuspended in lysis buffer (50 mM Tris-HCl, pH 7.5; 200 mM NaCl; 5% glycerol) supplemented with protease inhibitor cocktail. The cells were lysed by sonication, and the lysate was clarified by centrifugation. The supernatant was applied to a 5 mL HiTrap Q ion exchange column and eluted with a gradient up to 2 M NaCl. Peak fractions were pooled and adjusted to a final concentration of 3 M NaCl by mixing with 4 M NaCl buffer. This mixture was loaded onto a HiTrap Butyl HP column and eluted with a reverse gradient to 50 mM NaCl. Eluted peak fractions were concentrated to 5 mL using Vivaspin 15 10K MWCO filters and further purified by size exclusion chromatography (SEC) on a HiLoad 16/600 Superdex 75 pg column equilibrated with 20 mM Tris-HCl, pH 7.5, and 200 mM NaCl. The purified protein was concentrated, flash frozen, and stored at −80°C.
Purification of
bsu
ScpB
ScpB was purified following the protocol outlined by
25
. The coding sequence of ScpB, cloned into a pET-22 derived plasmid, was transformed into chemically competent BL21-Gold (DE3)
E. coli
cells. These cells were cultivated in ZYM-5052 autoinduction medium at 24°C for 23 hours. Subsequently, the cells were harvested and resuspended in lysis buffer (50 mM Tris-HCl, pH 7.5; 150 mM NaCl; 1 mM EDTA; 1 mM DTT) supplemented with protease inhibitor cocktail. Cell lysis was performed by sonication, followed by centrifugation to remove cell debris. The resulting supernatant was diluted to a final NaCl concentration of 50 mM and loaded onto a 5 mL HiTrap Q HP column. Elution was achieved using a gradient up to 2 M NaCl. The eluate was then diluted with lysis buffer containing 4 M NaCl to achieve a final concentration of 3 M NaCl. This sample was applied to two 5 mL HiTrap Butyl columns connected in series, and the protein was eluted with a reverse gradient to 50 mM NaCl. The peak fractions from this column were concentrated and subjected to size exclusion chromatography (SEC) using a HiLoad 16/600 Superdex 200 pg column equilibrated with 50 mM Tris-HCl, pH 7.5, 100 mM NaCl, and 1 mM DTT. Fractions containing ScpB were concentrated, flash frozen, and stored at −80°C.
Medium scale sybody purification
Individual sybody plasmids were transformed into chemically competent
E. coli
MC1061 cells via heat shock at 42 °C for 45 s, followed by recovery in LB medium at 37 °C. Transformed cells were cultured overnight in TB medium supplemented with 25 µg/ml chloramphenicol. Precultures were used to inoculate 50 mL TB cultures, which were grown at 37 °C before shifting to 22 °C. Expression was induced with 0.02% (wt/vol) L-(+)-arabinose and continued overnight at 22 °C with shaking. Cells were harvested by centrifugation and resuspended in periplasmic extraction buffer. Following incubation at 4 °C, cells were pelleted, and the supernatant was supplemented with imidazole to 15 mM. The extract was incubated with His MultiTrap HP resin, followed by centrifugation and washing with TBS containing 30 mM imidazole. Elution was performed using TBS with 300 mM imidazole. Purified sybodies were analyzed on a Sepax SRT-10C SEC100 column at 1 ml/min flow rate. Monomeric sybodies eluted between 11–12.5 ml, while retention volumes <11 ml indicated oligomerization, and >14 ml suggested strong column interaction. Non-expressed, oligomeric, or highly interacting sybodies were discarded. Typical yields ranged from 200 µg to 1 mg
16
.
Fluorescence imaging of ParB-GFP strains Image acquisition
B. subtilis
cells were first cultured in LB supplemented with 5 µg/mL chloramphenicol and 0.5% glucose at 30°C, from which a day culture was inoculated to 0.005 in LB with 5 µg/mL chloramphenicol. When needed, cultures were induced at OD
600
∼ 0.02 with 0.5% xylose and grown an extra 30 minutes until imaging at an OD
600
of 0.04. For microscopy analysis, 0.5 μL of the cell suspension was spotted onto agarose-coated microscopy slides. Images were acquired using a Leica DMi8 microscope equipped with an sCMOS DFC9000 (Leica) camera, a SOLA light engine (Lumencor), and a 100×/1.40 oil-immersion objective. Exposure time for image acquisition was set to 600 ms. Acquired images were processed using LAS X Office software (v.82, Leica Microsystems).
Image analysis
Microscopy images were analyzed using a fully automated image processing pipeline. Cell segmentation was performed with the pretrained Omnipose model
30
implemented in Cellpose
31
, which is optimized for bacterial morphologies. Prior to segmentation, a normalization step was applied to standardize image contrast. Following segmentation, intracellular foci were detected using Spotiflow
32
, a dedicated model trained for spot identification. Each detected spot was assigned to an individual cell based on spatial location, enabling quantification of the number of foci per cell. Cell morphology analysis was also performed to estimate cell length.
Quantitative results were compiled into a structured dataset, including the number of cells per image, percentage of cells lacking foci, number of foci per cell, cell length, and number of foci per micron. Data visualization and statistical analysis was performed using GraphPad Prism 10.4.1 (627).
ELISA
E. coli
MC1061 colonies were screened for sybody expression, and periplasmic extracts were obtained as per the original protocol
16
. ELISA plates were coated with Protein A and anti-c-Myc antibody before incubation with sybody-containing extracts. Three wells per sybody were incubated with biotinylated maltose binding protein (MBP, negative control) or with 50 nM biotinylated bsSmc-ScpAB(E1118Q) with ATP/dsDNA or without. Washes and ELISA development were performed using TBSM-D. Absorbance was measured at 650 nm.
ATPase measurements
ATPase activity was assessed using the coupled pyruvate kinase/lactate dehydrogenase reaction as described by
14
. ADP production was monitored over a period of 1 hour by measuring the absorbance changes of NADH at 340 nm. Data collection was performed using a Synergy Neo Hybrid Multi-Mode Microplate Reader. The reaction mixture comprised 1 mM NADH, 3 mM phosphoenolpyruvic acid, 100 U pyruvate kinase, 20 U lactate dehydrogenase, and varying concentrations of ATP. For assays requiring double-stranded oligonucleotides, a 40 bp oligonucleotide (5’-
TTAGTTGTTC
GTAGTGCTCG
TCTGGCTCTG
GATTACCCGC
-3’) was added to a final concentration of 3 mM. The final protein concentration in the assay was 0.15 μM
bsuS
mc dimers in ATPase assay buffer (50 mM HEPES-KOH, pH 7.5; 50 mM NaCl; 2 mM MgCl2). All measurements were conducted at 25°C.
Supplementary figures
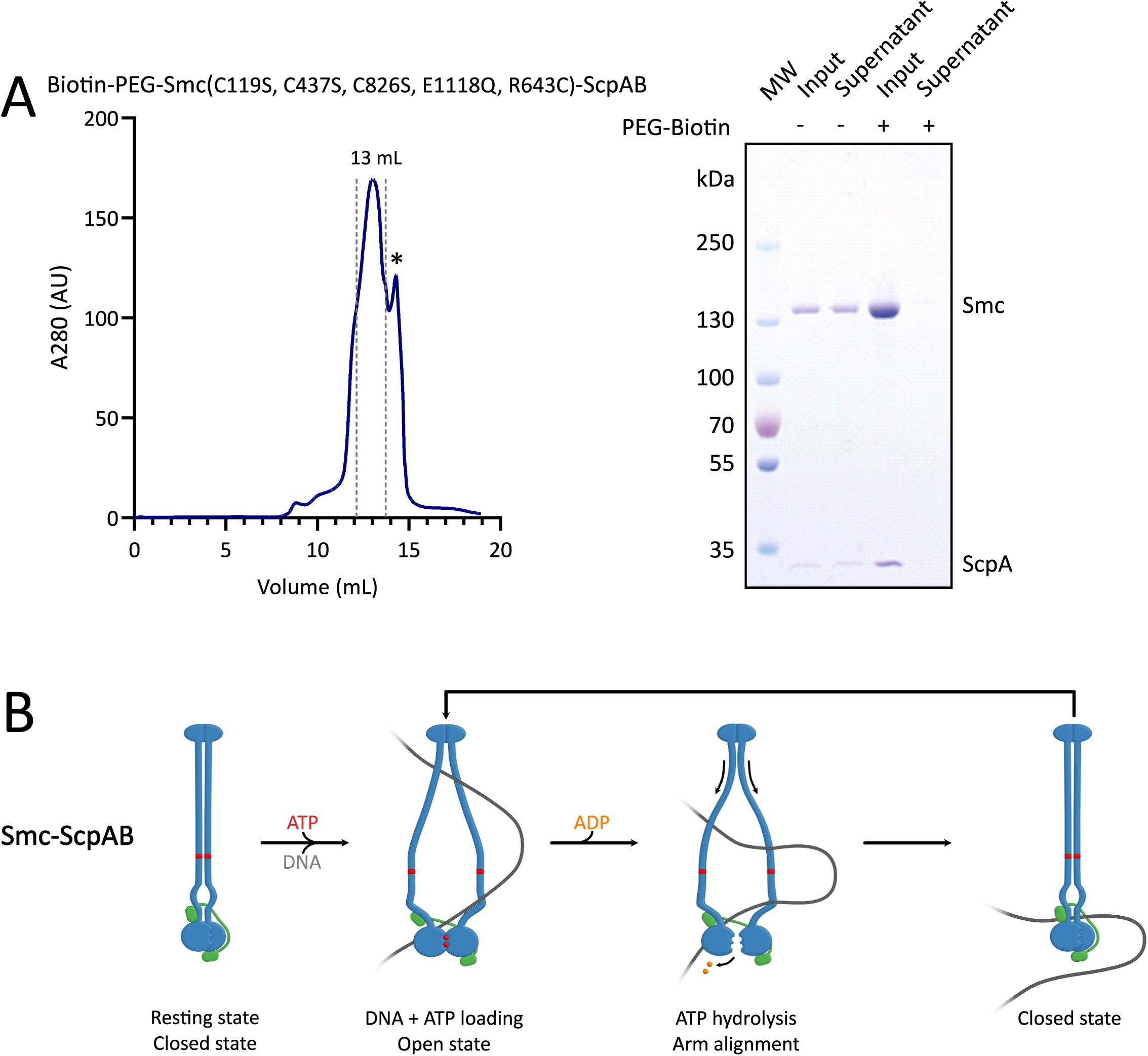
Preparation of
bsuS
mc(C119S, C437S, C826S, E1118Q, R643C)-ScpAB complex and loop extrusion model
(A)
Size-exclusion chromatography profile of biotinylated
bsuS
mc(C119S, C437S, C826S, E1118Q, R643C)-ScpAB complex. Peak fractions (elution at ∼13 mL, pooled fractions indicated by dotted lines for a final volume of 1.8 mL) were collected for downstream use; the final concentration was 5.9 µM (dimer). * : A secondary peak appeared at ∼14 mL; its identity was uncertain and was excluded from the peak. SDS-PAGE confirms successful biotinylation of the
bsu
Smc(C119S, C437S, C826S, E1118Q, R643C)-ScpAB complex.
(B)
Segment-capture model for
bsuS
mc-ScpAB, in which ATP binding and hydrolysis drive transitions between open and closed conformations to mediate loop extrusion.
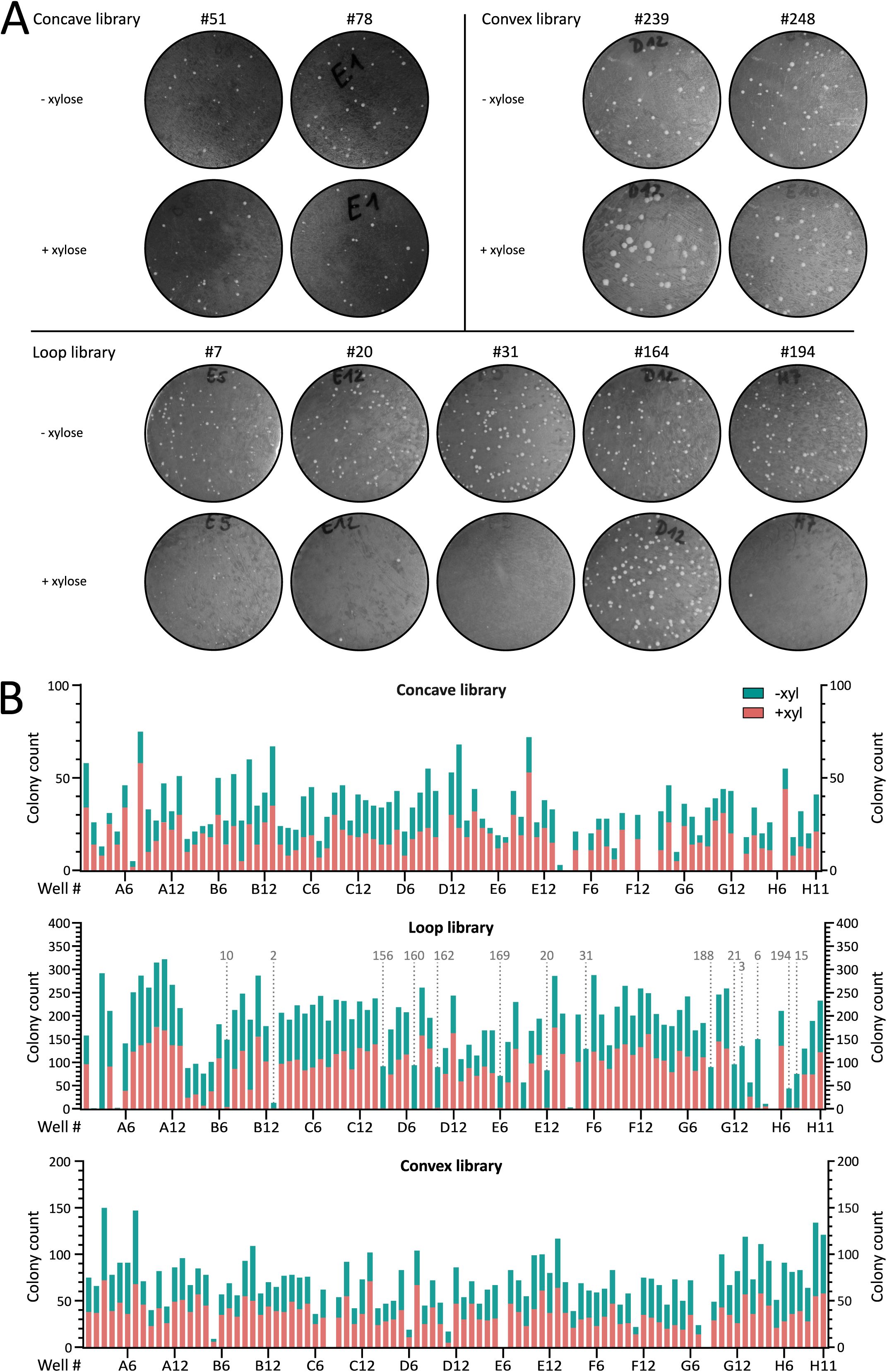
Representative results from
in vivo
sybody selection based on colony formation.
(A)
Representative examples of
in vivo
transformation assays for nine sybody-expressing
B. subtilis
strains. Sybody genes were integrated into the
amyE
locus under xylose-inducible control. Transformants were grown on oxoid nutrient agar (ONA) plates supplemented with chloramphenicol and 0.5% xylose to assess growth defects. Shown are candidates from the concave library: Sb051 (well B08) and Sb078 (D12). Loop library: Sb007 (E05), Sb020 (E12), Sb031 (F05), Sb164 (D12) and NbH07 (Sb194). Convex library: Sb239 (D12) and Sb248 (E10).
(B)
Growth assay results for the 285
B. subtilis
strains expressing individual xylose-inducible sybodies. Bars show each colony counts “without” (green) on top of “with” (pink) xylose. Strains are ordered by their original 96-well plate. Only the loop library yielded sybodies that consistently impaired growth under inducing conditions (dotted line). Sybody numbers indicated above the plots correspond to positive candidates used in subsequent experiments, numbered according to order of use. Notably, the strain corresponding to the E09 M sybody (Sb018), also showed an absence of growth upon sybody induction. However, this sybody candidate gave intermediate phenotypes in later experiments, which is why it was excluded from this study. Results for the loop library are also shown in
Figure 1
.
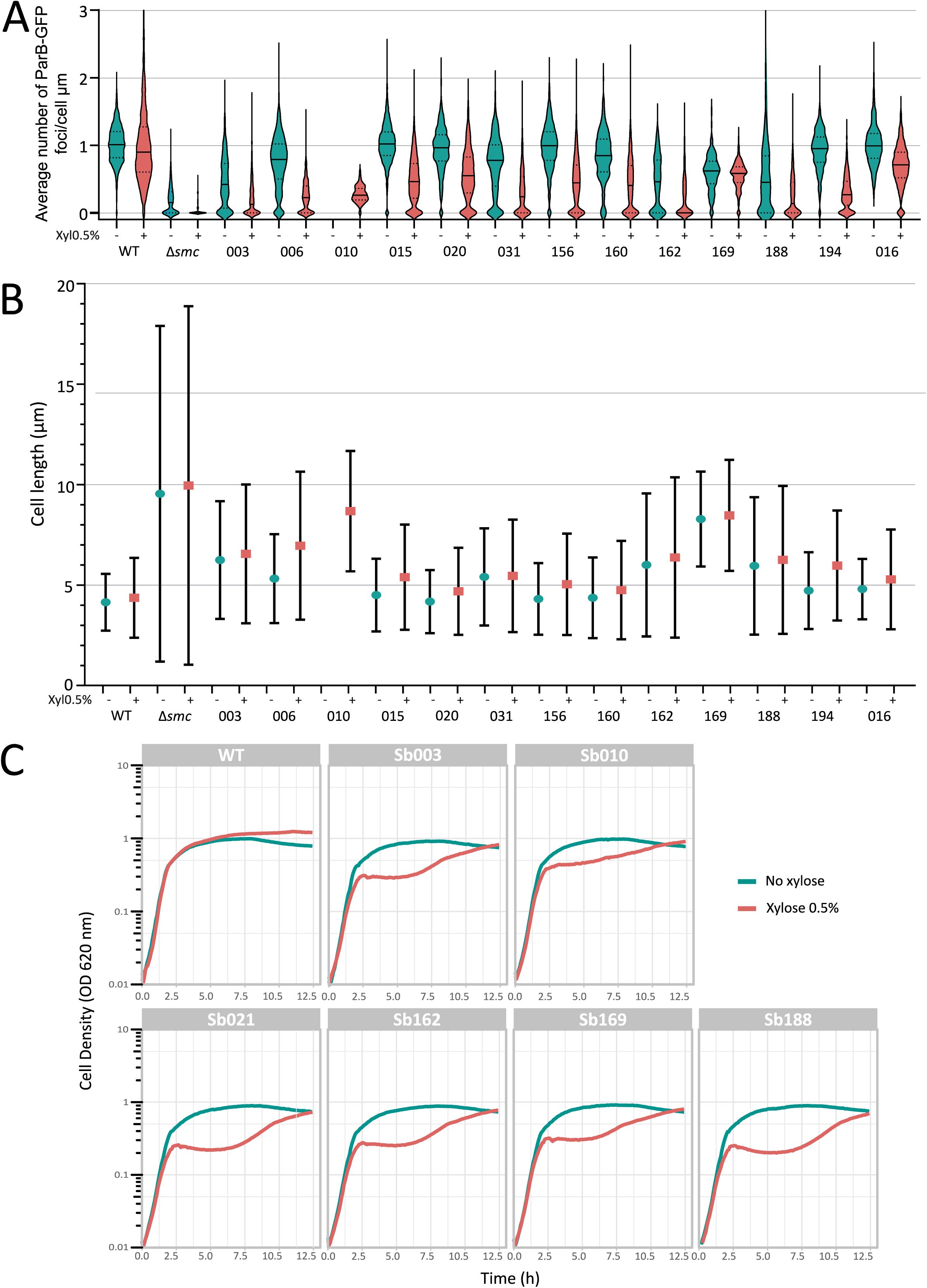
Functional impact of sybody expression on chromosome organization, cell length, and growth in
B. subtilis
.
(A)
Average number of ParB-GFP foci per µm of cell length in strains expressing individual sybodies, with and without xylose induction. Most uninduced strains differed significantly from the Smc WT control, likely due to leaky expression from the P
xyl
(
p
< 0.0001), except Sb015 (
p
> 0.9999), Sb156 (
p
= 0.2345), and Sb016 (negative control,
p
> 0.9999).
(B)
Average cell length across the same conditions, measured in ParB-GFP strains with or without Smc, and with or without each sybody. Bars indicate standard deviation. Tested sybodies caused cell elongation, a characteristic phenotype of impaired Smc activity, as chromosome segregation defects delay cell division, with mean lengths from 4.70 ± 2.16 µm (Sb020) to 8.68 ± 2.99 µm (Sb010), compared to 4.51 ± 1.95 µm in wild-type and 9.96 ± 8.92 µm in Δ
smc
.
(C)
Growth curves of
B. subtilis
strains expressing individual sybodies compared to the WT strain, with and without xylose induction. Each curve represents the mean of two biological replicates. Six sybodies were randomly picked and showed in this figure. Fluorophore-free strains showed wild type-like growth without xylose but strong delays post-induction, followed by partial growth recovery after 12 hours, likely due to xylose depletion or suppressor emergence. In sybody-expressing strains, a clear drop in cell density was observed ∼2.5 hours post-induction.

Sybody-GFP expression at different inducer concentration
(A)
Selected Sybody-GFP proteins were tested by GFP imaging at different inducer concentrations (0, 0.005, 0.05 and 0.5 % xylose). Representative images are shown. Notably, sybody Sb007 (E5, loop library) generates a mild growth phenotype (smaller colonies but normal colony numbers,
Figure 1A
and 1B
) but shows good expression levels and focal localization (top row).
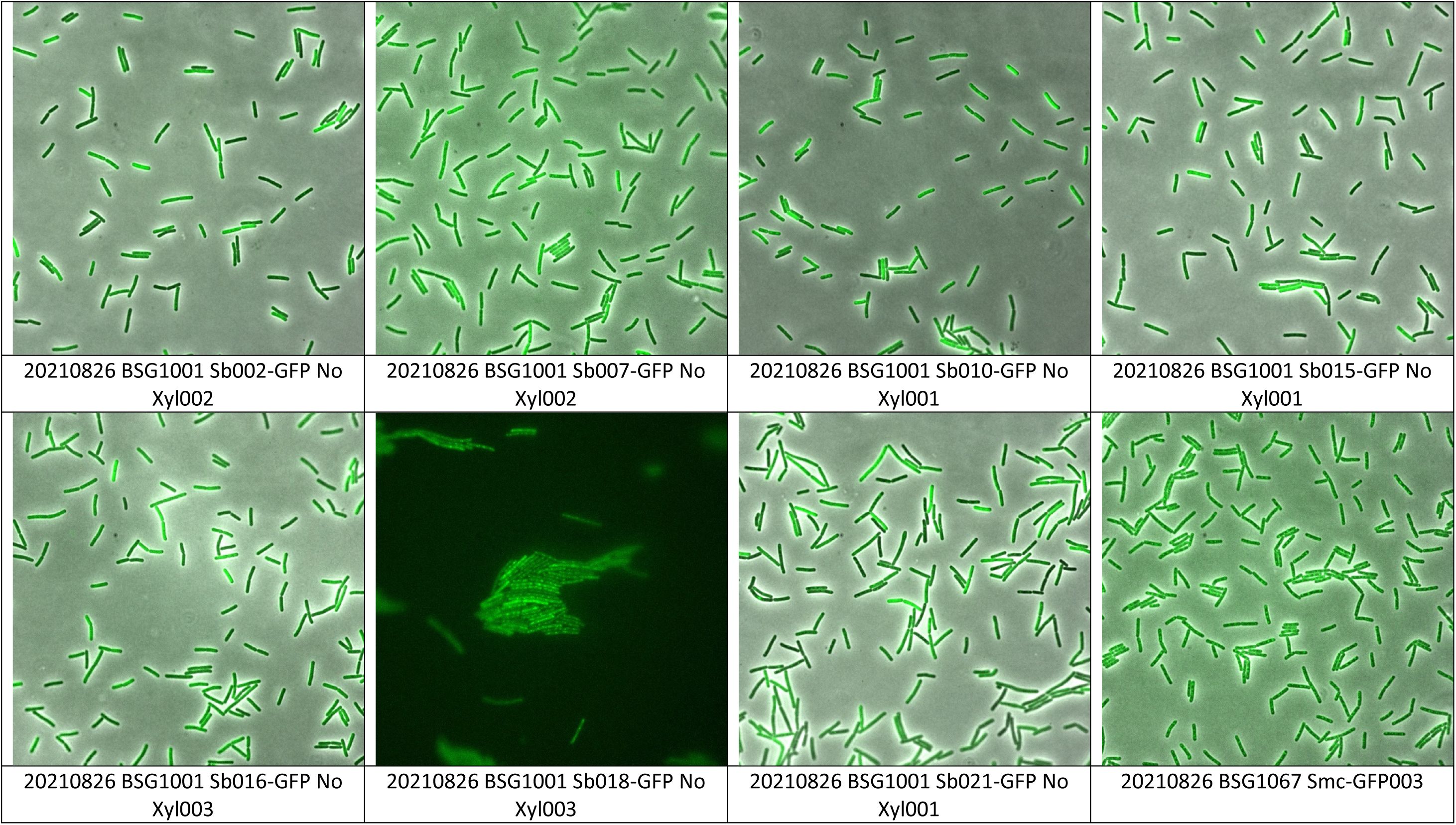
Imaging of various Sybody-GFP proteins without inducer.
(A)
Selected Sybody-GFP constructs were grown in the absence of inducer and tested for gfp expression. Representative images are shown.
ATPase hydrolysis rate of the
bsuS
mc-ScpAB complex with each selected sybody and viability of
B. subtilis
strains expressing chimeric Smc-ScpAB complexes in the absence of sybody induction.
(A)
Spotting assay assessing the growth of
B. subtilis
strains carrying chimeric Smc-ScpAB operons composed of
S. pneumoniae
and
B. subtilis
components. Strains harbor individual sybody constructs integrated at the
amyE
locus under control of a xylose-inducible promoter but were grown without inducer. “–” indicates no integration at
amyE
; “EV” corresponds to an empty vector control containing only the chloramphenicol resistance cassette. Numbered labels indicate strains carrying specific sybody constructs. Cells were spotted on rich medium (ONA) containing chloramphenicol and incubated for 16 h at 37 °C.
(B)
Overall ATPase hydrolysis rate of the
bsuS
mc-ScpAB complex in presence of each sybody and +/-DNA. Sb06, 021 and 194 triggered a significant difference in ATP hydrolysis between +/- dsDNA conditions. The rest is non-significant.
Hypothetical model for an Opening
DOWN
class of sybodies, not recovered in this study.
This may indicate that stabilizing an open conformation near the ATPase heads does not interfere with Smc function.
Data availability
All raw data will be made available at Mendeley Data DOI: 10.17632/k6k62p7z2s.1 (
).
Acknowledgements
We are grateful to Roberto Vazquez Nunez for the biotinylated
bsuS
mc(C119S, C437S, C826S, E1118Q, R643C) protein preparation, Arianna Ravera from the DCSR computing facility for help in image analysis and data extraction of the fluorescence microscopy images. Members of the Gruber Lab for critical feedback of the manuscript and for stimulating discussions.
Additional information
Funding
This study was supported by internal funding and by the European Research Council (Horizon 2020 ERC CoG 724482).
Author contributions
O.G. performed all
in vivo
experiments from sybody selection, fluorescence microscopy imaging, chimeric phenotypic screen. M.T. and L.H-H. performed the
in vitro
sybody screening (ribosome display, phage display, and first sybody purification). O.G. performed protein purifications, cross-linking and ATPase measurements, as well as data analysis, figures and writing. O.G. wrote the first manuscript draft. All authors reviewed and edited the manuscript. Funding acquisition and supervision, S.G.
Funding
EC | Horizon Europe | Excellent Science | HORIZON EUROPE European Research Council (ERC)
Stephan Gruber
Additional files
Table S1.
ELISA sybody binding assay.
Tables S4–S6.
Table S4.
Strains.
Table S5.
Plasmids.
Table S6.
Oligonucleotides.
References
1.
Synthetic single domain antibodies for the conformational trapping of membrane proteins
eLife
7
PubMed
Google Scholar
2.
One-shot design of functional protein binders with BindCraft
Nature
646
:483–492
PubMed
Google Scholar
3.
Organization of Chromosomal DNA by SMC Complexes
Annu Rev Genet
53
:445–482
PubMed
Google Scholar
4.
SMC condensin: promoting cohesion of replicon arms
Nat Struct Mol Biol
22
:653–655
PubMed
Google Scholar
5.
Recruitment of condensin to replication origin regions by ParB/SpoOJ promotes chromosome segregation in B. subtilis
Cell
137
:685–696
PubMed
Google Scholar
6.
The SMC condensin complex is required for origin segregation in Bacillus subtilis
Curr Biol
24
:287–292
PubMed
Google Scholar
7.
The SMC Hinge is a Selective Gate for Obstacle Bypass
Nat Commun
16
PubMed
Google Scholar
8.
Recruitment of SMC by ParB-parS organizes the origin region and promotes efficient chromosome segregation
Cell
137
:697–707
PubMed
Google Scholar
9.
Structure of Full-Length SMC and Rearrangements Required for Chromosome Organization
Mol Cell
67
:334–347
PubMed
Google Scholar
10.
DNA-segment-capture model for loop extrusion by structural maintenance of chromosome (SMC) protein complexes
Nucleic Acids Res
47
:6956–6972
PubMed
Google Scholar
11.
Control of Smc Coiled Coil Architecture by the ATPase Heads Facilitates Targeting to Chromosomal ParB/parS and Release onto Flanking DNA
Cell Rep
14
:2003–2016
PubMed
Google Scholar
12.
Structure of a nanobody-stabilized active state of the beta(2) adrenoceptor
Nature
469
:175–180
PubMed
Google Scholar
13.
Gradual opening of Smc arms in prokaryotic condensin
Cell Rep
35
:109051
PubMed
Google Scholar
14.
Tuned SMC Arms Drive Chromosomal Loading of Prokaryotic Condensin
Mol Cell
65
:861–872
PubMed
Google Scholar
15.
Positive and negative regulation of SMC-DNA interactions by ATP and accessory proteins
EMBO J
23
:2664–2673
PubMed
Google Scholar
16.
Generation of synthetic nanobodies against delicate proteins
Nat Protoc
15
:1707–1741
PubMed
Google Scholar
17.
Allosteric modulation of LRRC8 channels by targeting their cytoplasmic domains
Nat Commun
12
PubMed
Google Scholar
18.
High-Throughput Allelic Replacement Screening in Bacillus subtilis
Methods Mol Biol
2004
:49–61
PubMed
Google Scholar
19.
The extracellular gate shapes the energy profile of an ABC exporter
Nat Commun
10
:2260
PubMed
Google Scholar
20.
Interlinked sister chromosomes arise in the absence of condensin during fast replication in B. subtilis
Curr Biol
24
:293–298
PubMed
Google Scholar
21.
A joint-ParB interface promotes Smc DNA recruitment
Cell Rep
40
:111273
PubMed
Google Scholar
22.
Protein Sequence Analysis Using the MPI Bioinformatics Toolkit
Curr Protoc Bioinformatics
72
:e108
PubMed
Google Scholar
23.
Highly accurate protein structure prediction with AlphaFold
Nature
596
:583–589
PubMed
Google Scholar
24.
Molecular basis for SMC rod formation and its dissolution upon DNA binding
Mol Cell
57
:290–303
PubMed
Google Scholar
25.
Transient DNA Occupancy of the SMC Interarm Space in Prokaryotic Condensin
Mol Cell
75
:209–223
PubMed
Google Scholar
26.
Tuning the drug efflux activity of an ABC transporter in vivo by in vitro selected DARPin binders
PLoS One
7
:e37845
PubMed
Google Scholar
27.
Activation and allosteric modulation of a muscarinic acetylcholine receptor
Nature
504
:101–106
PubMed
Google Scholar
28.
An asymmetric SMC-kleisin bridge in prokaryotic condensin
Nat Struct Mol Biol
20
:371–379
PubMed
Google Scholar
29.
BactEXTRACT: an R Shiny app to quickly extract, plot and analyse bacterial growth and gene expression data
Access Microbiol
6
PubMed
Google Scholar
30.
Omnipose: a high-precision morphology-independent solution for bacterial cell segmentation
Nat Methods
19
:1438–1448
PubMed
Google Scholar
31.
Cellpose: a generalist algorithm for cellular segmentation
Nat Methods
18
:100–106
PubMed
Google Scholar
32.
Spotiflow: accurate and efficient spot detection for fluorescence microscopy with deep stereographic flow regression
Nat Methods
22
:1495–1504
PubMed
Google Scholar
Article and author information
Author information
Version history
Preprint posted
:
Sent for peer review
:
Reviewed Preprint version 1
:
Reviewed Preprint version 2
:
Cite all versions
You can cite all versions using the DOI
10.7554/eLife.111131
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2026,
Gosselin et al.
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
views
312
downloads
13
citations
0
Views, downloads and citations are aggregated across all versions of this paper published by eLife.





