 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库Recombination shapes the diversification of themeiotic drivers
Abstract
Meiotic drivers are selfish genetic elements that distort fair segregation. The
wtf
genes are poison-antidote meiotic drivers that are experiencing rapid diversification in fission yeasts. However, gene duplication alone is insufficient to drive the diversification of
wtf
genes, given the poison encoded by a newly duplicated
wtf
gene can be detoxified by the antidote encoded by the original
wtf
gene. Here, we analyze the evolution of
wtf
genes across 21 strains of
Schizosaccharomyces pombe
. Knocking out each of 25
wtf
genes in
S. pombe
strain 972h- separately does not attenuate the yeast growth, indicating that the
wtf
genes might be largely neutral to their carriers in asexual life cycle. Interestingly,
wtf
genes underwent recurrent and intricate recombination. As proof of principle, we generate a novel meiotic driver through artificial recombination between
wtf
drivers, and its encoded poison cannot be detoxified by the antidotes encoded by their parental
wtf
genes but can be detoxified by its own antidote. Therefore, we propose that recombination can generate new meiotic drivers and thus shape the diversification of the
wtf
drivers.
Introduction
During meiosis, the two alleles at a gene locus are separated into gametes, and each gamete has an equal chance of receiving either allele. This fundamental principle of inheritance, known as Mendel’s law of segregation (
Abbott and Fairbanks, 2016
), holds across most genetic loci in most sexual species. However, meiotic drivers, a class of selfish genetic elements, subvert fair segregation during gametogenesis and are transmitted to more than one-half (even to all) of the functional gametes produced by a heterozygote (
Sandler and Novitski, 1957
;
Lyttle, 1991
;
Hurst and Werren, 2001
;
Bravo Núñez et al., 2018b
). Meiotic drivers can spread in a population even when they impose fitness costs on their hosts (
Crow, 1991
;
Lindholm et al., 2016
). However, the spread of a meiotic driver can be thwarted by the costs imposed on its carriers or by its genetic suppressors (
Lindholm et al., 2016
).
The fission yeast
wtf
(
w
ith
Tf
Long Terminal Repeats
) gene family provides an excellent model to study how meiotic drivers act and evolve (
Hu et al., 2017
;
Nuckolls et al., 2017
). Many
wtf
genes are autonomous one-gene poison-antidote meiotic drivers that encode both a spore-killing poison (short isoform) and an antidote to the poison (long isoform) using alternative transcriptional initiation (
Hu et al., 2017
;
Nuckolls et al., 2017
;
Nuckolls et al., 2020a
). To achieve meiotic drive, all spores are exposed to the poison, whereas only those that inherit
wtf
express the antidote and are rescued (
Hu et al., 2017
;
Nuckolls et al., 2017
;
Nuckolls et al., 2020b
). Some other
wtf
genes can act as drive suppressors (
Bravo Núñez et al., 2018a
;
Bravo Núñez et al., 2020a
). The poison and the antidote differ only in their N-terminal cytosolic tails containing PY (Leu/Pro-Pro-X-Tyr) motifs. PY motif-dependent ubiquitination promotes the transport of the antidote and the poison (physically interacted with the antidote) from the trans-Golgi network to the endosome, thereby preventing toxicity (
Zheng et al., 2023
).
The
wtf
gene family is experiencing rapid diversification: the
Schizosaccharomyces pombe
reference genome encodes 25
wtf
genes, some of which are pseudogenes. The copy numbers of
wtf
genes vary greatly among different
S. pombe
strains, and frequent nonallelic gene conversion occurs between
wtf
genes (
Hu et al., 2017
;
Nuckolls et al., 2017
;
Eickbush et al., 2019
). However, these findings are based on a limited number of strains, and the pattern and extent of recombination in the
wtf
genes remain to be fully explored. Moreover,
wtf
driver genes are present in the last common ancestor (LCA) of the fission yeasts
S. pombe
,
S. octosporus
,
S. osmophilus
, and
S. cryophilus
, indicating that
wtf
genes have likely maintained the capacity to drive for more than 100 million years (
De Carvalho et al., 2022
). These fission yeast species carry varying numbers of
wtf
genes, ranging from 5 to 83 (
De Carvalho et al., 2022
). Yet, it remains perplexing how
wtf
genes achieved such diversification. On one hand, gene duplication can give birth to new
wtf
gene copies. On the other hand, a newly duplicated
wtf
gene might not drive because the poison produced by the newly duplicated
wtf
gene can be detoxified by the original
wtf
gene. Like newly duplicated genes, the most probable fate of a new
wtf
duplicate is pseudogenization (
Lynch, 2007
;
Innan and Kondrashov, 2010
). Thus, the vast majority of new
wtf
duplicates experience an early exit from the population, most probably never reaching fixation (
Lynch, 2007
;
Innan and Kondrashov, 2010
). To become a new driver, the new
wtf
copy should evolve coupling new poison and new antidote to the new poison through mutations. But the fate-changing mutations are likely to be rare. It follows that gene duplication might be insufficient to drive the diversification of
wtf
genes.
In this study, we analyzed the diversity and evolution of
wtf
genes in the genomes of 21 strains of
S. pombe
that were sequenced using long-read sequencing approaches (
Tusso et al., 2022
). Through knocking out each of 25
wtf
genes in
S. pombe
laboratory strain 972h-, no significant attenuated growth was observed, indicating
wtf
genes might be not deleterious in the asexual life cycle. We found that recurrent recombination occurred among
wtf
genes. We generated a novel meiotic driver through artificial recombination between
wtf
drivers, and its encoded poison cannot be detoxified by the antidotes encoded by their parental
wtf
genes but can be detoxified by its own antidote. Therefore, we propose that recombination can generate
wtf
driver with new poisons and might shape the diversification of
wtf
genes.
Results
Diversity and evolution of
wtf
genes in fission yeasts
First, we analyzed the diversity and distribution of
wtf
genes in fission yeasts. The
S. pombe
reference genome (strain 972h-) encodes a total of 25
wtf
genes. For these 25
wtf
genes, the number of exons varies from 3 to 6 (
Figure 1A
;
Bowen et al., 2003
). To investigate the relationship among exons from different
wtf
genes, we grouped these
wtf
exons into clusters based on nucleotide identity of 0.50 (
Figure 1B and C
). The
wtf
exons were grouped into 10 clusters with >2 members, and 12 exons exist as singletons in the similarity network (
Figure 1B and C
,
Supplementary file 1a
). Only exon 1 of all the
25 wtf
genes group together in a cluster, indicating that the first exons are well conserved among the
wtf
genes. No other exon is conserved among all the 25
wtf
genes. Therefore, the evolution of the
wtf
gene structures appears to be highly dynamic.
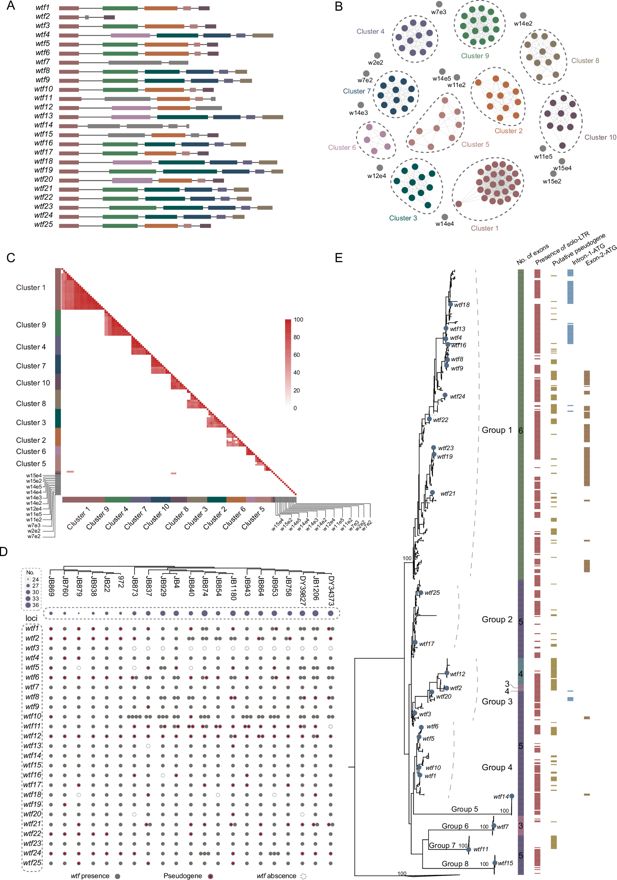
The diversity and evolution of
wtf
genes in fission yeasts.
(
A
) The gene structures of the
wtf
genes in the
S. pombe
reference genome. Rectangles and lines represent exons and introns, respectively. Rectangles with the same color indicate exons that share >50% sequence identity. (
B
) The similarity network of exons of the
wtf
genes in the
S. pombe
reference genome. Sequences that share an identity of >50% form a cluster. The colors of
wtf
exons correspond to these in panel (
A
). The exons for each cluster were listed in the
Supplementary file 1a
. (
C
) The heatmap of nucleotide identity among exons of the
wtf
genes in
S. pombe
reference genome. (
D
) The distribution of
wtf
genes in 21
S. pombe
strains.
wtf
genes were present in 25 genetic loci. Filled circles and empty circles represent the presence or absence of
wtf
genes, respectively. Red pentagons indicate putative
wtf
pseudogenes. The size of circles in purple indicates the number of
wtf
genes. The relationship among 21
S. pombe
strains was inferred based on phylogenetic analysis of 30 randomly selected genes. (
E
) Phylogenetic relationship of
wtf
genes in 21
S. pombe
strains. The phylogenetic tree is reconstructed using the maximum likelihood method. Filled circles in blue indicate
wtf
genes in
S. pombe
reference genome. The number of exons, the presence of solo-LTRs, the putative pseudogene status, the presence of intron-1-ATG codon, and the presence of exon-2-ATG codon are shown near the corresponding
wtf
gene.
wtf
genes from other fission yeast species were collapsed into a triangle.
Figure 1—source data 1
Gene structures for wtf1 to wtf25.
Download elife-100638-fig1-data1-v1.xlsx
Figure 1—source data 2
The sequence identity among all the exons of wtf elements.
Download elife-100638-fig1-data2-v1.xlsx
Figure 1—source data 3
The distribution and information of wtf genes in 21
S. pombe
strains.
Download elife-100638-fig1-data3-v1.xlsx
We next identified
wtf
genes in 21 strains of
S. pombe
that were sequenced using long-read sequencing approaches (
Supplementary file 1b
;
Tusso et al., 2022
). The copy number of
wtf
genes varies among different
S. pombe
strains, ranging from 24 (strain JB879) to 37 (strain JB1206) (
Figure 1D
). Synteny analyses show that the
wtf
genes are present in 20 genetic loci (
Figure 1D
). Multiple
wtf
genes were present in 13
wtf
loci. Within 20
wtf
loci, at least one
wtf
gene is present in all of or nearly all of the 21
S. pombe
strains, suggesting that these 20
wtf
loci might have originated before the LCA of the 21
S. pombe
strains.
wtf
pseudogenes are prevalent in many
wtf
loci among the 21
S. pombe
strains, indicating frequent pseudogenization occurred in the
wtf
genes. These results indicate that
wtf
copy number variation is prevalent among
S. pombe
strains.
We performed phylogenetic analyses of the
wtf
genes from 21
S. pombe
strains and three other fission yeast species (
S. octosporus
,
S. cryophilus
, and
S. osmophilus
) (
Figure 1E
,
Supplementary file 1c
). The
wtf
genes of
S. pombe
form a monophyletic group. Orthologs of
wtf14
,
wtf7
,
wtf11
, and
wtf15
form monophyletic groups, whereas orthologs of other
wtf
genes show complex phylogenetic mixing, indicating complex recombination might have occurred among these
wtf
genes (
Figure 1E
;
Eickbush et al., 2019
). Moreover, the
wtf
genes with six exons (including the known meiotic drivers
wtf4
,
wtf9
,
wtf13,
and
wtf23
) (
Nuckolls et al., 2017
;
Bravo Núñez et al., 2018a
;
Bravo Núñez et al., 2020a
) cluster together and exhibit a ladder-like phylogeny, which might be generated by continual selection driven by antidotes (like the ladder-like phylogeny of influenza A viruses H1N1 and H3N2, which is shaped by continual immune selection;
Grenfell et al., 2004
;
Bedford et al., 2011
). Based on phylogenetic relationship, we divided the
wtf
genes of 21
S. pombe
strains into eight groups, namely groups 1–8, among which groups 5–8 include orthologs of
wtf14
,
wtf7
,
wtf11
, and
wtf15
, respectively (
Figure 1E
). Exon 2 ATG codons (exon-2-ATG) and in-frame ATG within intron 1 and near the start of exon 2 (intron-1-ATG) of
wtf
genes can encode the start of poison protein isoforms (
Hu et al., 2017
). We found that most of exon-2-ATG and intron-1-ATG are present within group 1
wtf
genes (
Figure 1E
). A majority of the
wtf
genes are flanked by solo-LTRs (
Bowen et al., 2003
;
Figure 1E
). However, the solo-LTRs flanking the
wtf
genes do not cluster together but form many distinct groups, suggesting that solo-LTRs were inserted nearby the
wtf
genes multiple times (
Figure 1—figure supplement 1
). Together, our results reveal the rapid diversification and turnover of
wtf
genes in a single fission yeast species.
No attenuated growth of fission yeast without
wtf
genes
To explore the effect of
wtf
genes on the fitness of fission yeast, we knocked out each of the 25
wtf
genes in the
S. pombe
laboratory strain 972h- using a method based on homologous recombination (
Figure 2A
). A total of 25
wtf
knockout strains (
Δwtf1
to
Δwtf25
) were generated. We used spot assay to evaluate the effect of
wtf
gene knockout on the yeast growth, and no growth defect was observed for all the 25
wtf
knockout strains (
Figure 2B
). Furthermore, no significant differences were observed in the growth curves between the wild-type and
wtf
knockout strains (
Figure 2C
) or in the maximum growth rates among the wild-type and
wtf
knockout strains. Therefore, our experiment suggests that the
wtf
genes might be largely neutral to the fitness of their carriers in the asexual life cycle at least in normal growth condition.
Figure 2
Download asset
Open asset
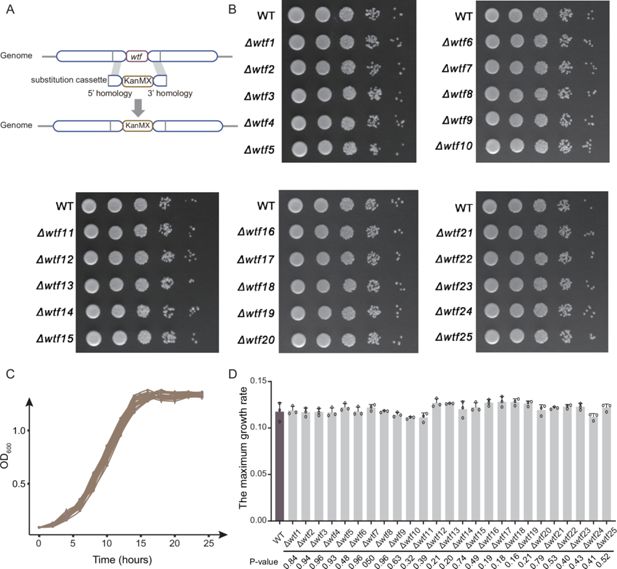
The effect of
wtf
gene on the growth of
S. pombe
.
(
A
) Generation of
wtf
knockout (
Δwtf
) strains based on the homologous recombination method. Substitution cassette contains a kanMX resistance marker and two homologous sequences flanking the target
wtf
genes. (
B
) Spot assay of
Δwtf
strains. The strains were diluted in five 10-fold steps to 10
–5
, and 1.5 μL of each dilution were spotted on the surface of YE solid media. Growth curves (
C
) and maximum growth rates (
D
) of WT and 25
wtf
knockout strains. Data represent means of three biological replicates (solid lines or bars), with error bars showing SD. Open circles indicate individual replicate values.
Figure 2—source data 1
Data for growth rates of wtf knockout strains.
Download elife-100638-fig2-data1-v1.xlsx
Recurrent recombination in
wtf
genes
Given complex phylogenetic mixing observed among
wtf
genes (
Figure 1E
), we tested whether recombination occurred. We detected signals of recombination in the 25
wtf
genes of the
S. pombe
reference genome (p<0.0001) and in the
wtf
genes of the 21
S. pombe
strains (p<0.0001) using pairwise homoplasy index (HPI) test. Split network analysis also supports the frequent occurrence of recombination in the 25
wtf
genes of the
S. pombe
reference genome (
Figure 3A
) and in the
wtf
genes of the 21 strains of
S. pombe
(
Figure 3B
). In contrast, no recombination signal was detected for groups 5–8 using HPI test (p=1 for group 5, p=1 for group 6, p=0.53 for group 7, and p=1 for group 8). We estimated recombination rates of the full-length
wtf
sequences, the first exons, and the
wtf
sequences without the first exons for
wtf
groups 1–4. We found that the recombination rate of group 1
wtf
was highest among the four
wtf
groups (
Figure 3C
). For group 1, breakpoints are dispersed across the
wtf
sequences (
Figure 3D
). These lines of evidence suggest that
wtf
genes underwent recurrent and intricate recombination.
Figure 3
Download asset
Open asset
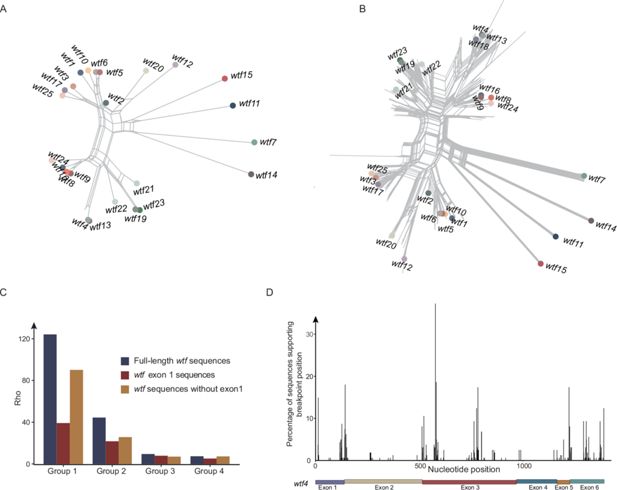
Recombination analysis of
wtf
genes.
(
A
) Split tree of 25
wtf
genes of the
S. pombe
reference genome. (
B
) Split tree of
wtf
genes from 21
S. pombe
strains. The
wtf
genes of the
S. pombe
reference genome are labeled. (
C
) Recombination rates of
wtf
genes that belong to groups 1–4. Recombination rates were estimated for the full-length
wtf
sequences, the first exons, and the
wtf
sequences without the first exons. (
D
) Breakpoints detected for
wtf
genes of group 1. The exons of
wtf4
, as a gene position reference, are shown.
Generation of a new driver gene through artificial recombination
Given gene duplication alone might be insufficient to shape the diversification of
wtf
genes, we hypothesize that recombination between
wtf
genes can generate new meiotic drives. To test this, we constructed four chimeric
wtf
genes through recombination among known functional meiotic drivers (
wtf23
) (
Bravo Núñez et al., 2018a
;
Bravo Núñez et al., 2020a
) and an artificially generated meiotic driver (
wtf18
) as specified below.
We used a proved
Saccharomyces cerevisiae
system to test the activity of poison and antidote proteins encoded by
wtf
genes (
Nuckolls et al., 2020a
). As expected, the expression of the poison proteins (Wtf23
poison
) encoded by
wtf23
genes caused the yeast growth arrest (
Figure 4A
). The attenuated growth was alleviated when the corresponding antidote proteins (Wtf23
antidote
) were expressed (
Figure 4A
). We also experimentally analyzed
wtf18
gene, which was known to encode only long (antidote-like) transcripts and probably act as a suppressor (
Bravo Núñez et al., 2018a
). We artificially introduced an in-frame ATG codon right before the start of exon 2, generating
wtf18
poison/-0M
. The expression of
wtf18
poison/-0M
resulted in the yeast growth arrest, suggesting its product, Wtf18
poison/-0M
, is indeed a poison protein (
Figure 4B
). When co-expressing
wtf18
antidote
and
wtf18
poison/-0M
, the attenuated yeast growth was rescued (
Figure 4B
), indicating that Wtf18
antidote
can ameliorate the toxicity of Wtf18
poison/-0M
.
Figure 4
Download asset
Open asset
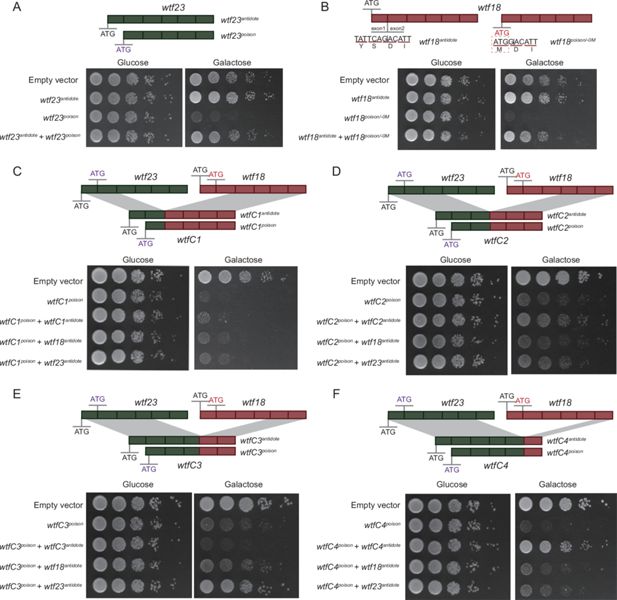
Poison and antidote activity of
wtf
genes and chimeric
wtf
genes.
wtf23
and
wtf18
are highlighted in green and red, respectively. Rectangles represent exons. The start codon ATG is shown. For
wtf18
, an in-frame ATG codon was introduced right before the start of exon 2, generating
wtf18
poison/-0M
. Spot assay of yeast transformed with the short isoforms (encoding poison-like proteins) and the long isoforms (encoding antidote-like proteins) of
wtf23
(
A
) and
wtf18
(
B
). (
C–F
) Four chimeric
wtf
genes were generated through artificial recombination, namely
wtfC1
(with exons 1–2 of
wtf23
and exons 3–6 of
wtf18
),
wtfC2
(with exons 1–3 of
wtf23
and exons 4–6 of
wtf18
),
wtfC3
(with exons 1–4 of
wtf23
and exons 5–6 of
wtf18
), and
wtfC4
(with exons 1–5 of
wtf23
and exon 6 of
wtf18
). Spot assay of yeast transformed with the short isoforms (encoding poison-like proteins) and the long isoforms (encoding antidote-like proteins) of
wtfC1
(
C
),
wtfC2
(
D
),
wtfC3
(
E
), and
wtfC4
(
F
) are shown.
We then constructed four chimeric
wtf
genes through artificial recombination between
wtf23
and
wtf18
, including
wtfC1
(possessing exons 1–2 of
wtf23
and exons 3–6 of
wtf18
),
wtfC2
(possessing exons 1–3 of
wtf23
and exons 4–6 of
wtf18
),
wtfC3
(possessing exons 1–4 of
wtf23
and exons 5–6 of
wtf18
), and
wtfC4
(possessing exons 1–5 of
wtf23
and exon 6 of
wtf18
). The expression of the short isoforms of
wtfC1
,
wtfC2
,
wtfC3
, and
wtfC4
resulted in yeast growth arrest, revealing their toxicity (
Figure 4C–F
). However, the antidote of
wtfC1
and
wtfC3
cannot detoxify the corresponding chimeric toxins (
Figure 4C and E
). Interestingly, we generated a putative novel meiotic driver, namely
wtfC4
. Our results show that
wtfC4
encodes a functional poison (WtfC4
poison
) (
Figure 4F
). The poison can be detoxified by its own long isoforms (dubbed as
wtfC4
antidote
), but cannot be detoxified by the antidote proteins of their parental genes (
Figure 4F
). Taken together, we generated a new meiotic driver through artificial recombination between pre-existing
wtf
genes.
We tried to test the driver phenotype of
wtfC4
in a more natural setting. We created a recombinant strain,
Sp-wtfC4
, based on the laboratory strain 972h-. Specifically, we replaced the last exon of the original
wtf23
gene with the last exon of
wtf18
. However, we encountered a challenge: since strain 972h- has only one mating type and cannot undergo meiosis on its own, we had to mate the recombinant strain with a BN0 h
+
strain that only carries
wtf23
antidote
. We did not observe the meiotic driver phenotype as expected. This might be due to issues with the proper splicing and expression of the potential poison and antidote proteins or due to the genetic background. Nevertheless, our results raise the possibility that new meiotic drivers can arise through recombination.
Discussion
In this study, we analyzed the diversity and evolution of
wtf
genes in fission yeasts. The copy number of the
wtf
gene varies among different
S. pombe
strains, revealing rapid diversification and turnover of the
wtf
genes within a single fission yeast species. We detected signals of recurrent and intricate recombination among
wtf
genes as previously reported with limited genomes (
Hu et al., 2017
;
Nuckolls et al., 2017
;
Eickbush et al., 2019
;
De Carvalho et al., 2022
). We hypothesize that recombination between
wtf
genes can produce new
wtf
genes with new poisons and the antidotes to new poisons. These new
wtf
genes can then drive through populations. As proof of principle, we generated a chimeric
wtf
gene that represents a new meiotic driver. The encoded poison of the newly generated meiotic driver can be detoxified by its own long isoforms, but cannot be detoxified by the antidote proteins of their parental genes. However, the other three chimeric
wtf
genes tested did not show this property. Indeed, our recombination breakpoint analyses (
Figure 3D
) reveal substantial recombination might have occurred in the last exon. Together, our results indicate that recombination is likely to drive the rapid diversification of
wtf
gene in fission yeasts.
Most of the known meiotic drivers impose costs on their carriers due to direct effects of the driver on survival or fertility, production of a biased sex ratio, or via deleterious mutations linked to the driver (
Price and Wedell, 2008
;
Larracuente and Presgraves, 2012
;
Sutter and Lindholm, 2015
;
Fishman and Kelly, 2015
;
Lindholm et al., 2016
;
Zanders and Unckless, 2019
). In outcrossing between individuals from distinct yeast lineages,
wtf
drivers can provide a selective advantage to atypical spores, such as aneuploids and diploids (
Bravo Núñez et al., 2020b
). In this study, we assessed the effects of the
wtf
genes on the growth of fission yeast during the asexual life cycle through knocking out each of the 25
wtf
genes in
S. pombe
laboratory strain 972h- separately. We did not observe obvious attenuated growth for these
wtf
knockout strains, indicating
wtf
genes are largely neutral to the fitness of their carriers during the asexual life cycle at least in the normal growth setting. It should be noted that the spot assay used in this study detects only large differences in fitness between wild type and
wtf
knockout strains. Nevertheless, it is likely that
wtf
genes evolve mainly in a neutral manner during the asexual life cycle, which explains the presence of a high proportion of pseudogenes in
wtf
gene repertoire. Moreover, asexual reproduction is much more frequent than sexual reproduction for yeasts (
Tsai et al., 2008
). Therefore, even if fate-changing mutations that simultaneously produce new poison and the antidote to new poison occur, the most probable fate of a new
wtf
gene generated by gene duplication is pseudogenization and removal from the population.
Gene duplication gives rise to new
wtf
genes. However, the newly generated
wtf
gene can be detoxified by the original
wtf
gene and thus cannot drive through its host population when the original
wtf
is fixed in the population (
Figure 5
). Therefore, most, if not all, of the
wtf
gene duplicates experience early exit from the host population. When recombination occurs between two pre-existing
wtf
genes, chimeric
wtf
gene with new poison and the antidote to new poison can be generated as this study shows. Then, the
wtf
gene with new driver property can spread in its host population, even reaching fixation (
Figure 5
). During asexual life cycle,
wtf
genes evolve mainly under genetic drift, and thus can accumulate disruptive mutations, leading to their pseudogenization. Taken together, our study highlights the significance of recombination in shaping the diversification of
wtf
genes.
Figure 5
Download asset
Open asset
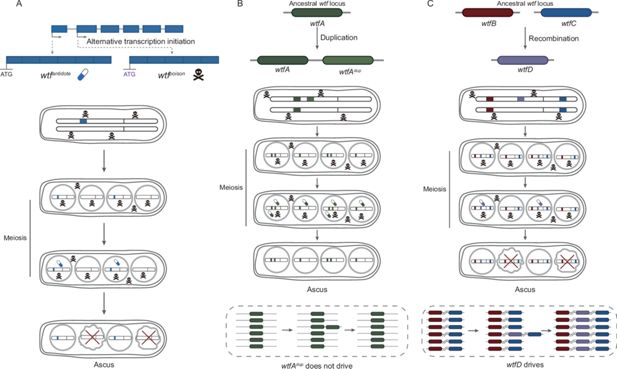
The evolutionary mechanisms of
wtf
genes.
(
A
) Mechanism of
wtf
meiotic driver.
wtf
genes are transcribed into two transcripts, namely long and short isoforms. The long and short isoforms encode antidote and poison, respectively. All spores are exposed to the poison, whereas only those that inherit
wtf
genes express the antidote and are rescued. Spores without the
wtf
allele are destroyed, resulting in the drive of
wtf
. (
B
) Duplication is insufficient to drive the diversification of
wtf
genes. Gene duplication of
wtfA
gives rise to a new
wtf
gene,
wtfA
dup
. However,
wtfA
dup
gene can be detoxified by the original
wtfA
gene and thus cannot drive through its host population. (
C
) When recombination occurs between two parental
wtfB
and
wtfC
genes, generating chimeric
wtfD
gene.
wtfD
encodes a new poison that cannot be detoxified by their parental antidote and the antidote to the new poison. The
wtfD
gene then can spread in its host population.
Methods
Identification of the
wtf
genes
We used the blastn algorithm to identify
wtf
genes within 21
S. pombe
strains with 25
wtf
genes from
S. pombe
reference genome as the queries and an
e
-cutoff value of 10
–5
. The identified
wtf
genes were annotated based on the
wtf
genes of the reference genome. The sequence identity among the exons of
wtf
genes was calculated using BioAider version 1.334 (
Zhou et al., 2020
). Exons were then clustered based on the nucleotide identity using the igraph package version (
Csardi and Nepusz, 2006
;
Csárdi et al., 2024
). We extended 1000 bp flanking each
wtf
gene to establish their syntenic relationships.
Phylogenetic analysis
The coding sequences of
wtf
genes of 21
S. pombe
strains and three other fission yeast species (
S. octosporus
,
S. cryophilus
, and
S. osmophilus
) were aligned using MAFFT version 7 (
Katoh and Standley, 2013
). To clarify the relationship of 21
S. pombe
strains, 30 genes were randomly selected and concatenated using Phylosuite version 1.2.1 (
Zhang et al., 2020
). All the phylogenetic analyses in this study were performed using the maximum likelihood (ML) method implemented in IQ-TREE version 2 (
Minh et al., 2020
). The best-fit substitution model was selected using the ModelFinder algorithm (
Kalyaanamoorthy et al., 2017
). Node supports were assessed using the ultrafast bootstrap approximation (UFBoot) method with 1,000 replicates (
Hoang et al., 2018
). Solo-LTRs were identified using the blast algorithm and aligned using MAFFT version 7 (
Katoh and Standley, 2013
). Phylogenetic analysis was performed using the approximate maximum likelihood method implemented in FastTree version 2.1.1 (
Price et al., 2010
).
Recombination analysis
Split networks of
wtf
genes were generated using the neighborhood network analysis implemented in SplitsTree4 (
Huson and Bryant, 2006
). Pairwise homoplasy index (PHI) test was performed using SplitsTree4 (
Huson and Bryant, 2006
). Potential breakpoints were detected using 3SEQ (
Lam et al., 2018
). The recombination rate was estimated using the FastEPRR package version 2.0 (
Gao et al., 2016
). The recombination rate was estimated using FastEPRR version 2.0 (
Gao et al., 2016
) as the population-scaled recombination rate, Rho = 4Nₑr, where Nₑ is the effective population size and r is the per-generation recombination rate. This scaling allows comparison of recombination rates across genomic regions and populations.
Generation of
wtf
knockout strains
The
wtf
knockout (
Δwtf
) strains generated in this study were derived from
S. pombe
strain 972h-. We constructed substitution cassettes for each of the 25
wtf
genes of the
S. pombe
reference genome. Substitution cassettes contain a kanMX resistance marker and two homologous sequences flanking the target
wtf
genes (
Moreno et al., 1991
;
García-Ríos et al., 2014
). Substitution cassettes were transformed into
S. pombe
(strain 972h-) through the lithium acetate-based method (
Moreno et al., 1991
;
García-Ríos et al., 2014
).
wtf
gene knockout strains were selected for kanMX resistance and were verified by PCR. The primers used for
wtf
knockout in
S. pombe
are provided in
Supplementary file 1d
.
Plasmid construction
Total RNA of fission yeast was extracted and reverse transcribed into cDNA. Coding sequences of
wtf23
antidote
,
wtf23
poison
, and
wtf18
antidote
were amplified using the corresponding primers (
Supplementary file 1e
).
wtf18
poison/-M0
was generated using
wtf18
antidote
as the template and the primer with an artificially introduced ATG (
Supplementary file 1e
). We generated
wtfC1
through recombining exons 1–2 of
wtf23
and exons 3–6 of
wtf18
, generated
wtfC2
through recombining exons 1–3 of
wtf23
and exons 4–6 of
wtf18
, generated
wtfC3
through recombining exons 1–4 of
wtf23
and exons 5–6 of
wtf18
, and generated
wtfC4
through recombining exons 1–5 of
wtf23
and exon 6 of
wtf18
. These
wtf
and
wtfC
genes were then cloned into the GAL1/10 dual expression plasmid Gal_HF. Plasmids were first transformed into
Escherichia coli
and verified by PCR and sequencing. Plasmids were then transformed into
S. cerevisiae
(strain S288C) using the lithium acetate-based method. Yeast transformants were selected for kanMX resistance and were verified by PCR.
Meiotic analysis
We created a recombinant strain,
Sp-wtfC4
, based on the laboratory strain 972h-. Specifically, we replaced the last exon of the original
wtf23
gene with the last exon of
wtf18
using homologous recombination. The
SP-wtfC4
strain and the BN0 h
+
strain carrying
wtf23
antidote
were streaked separately onto YE solid plates and incubated at 30°C for approximately 20 hours. Cells from each strain were then scraped and resuspended in sterile water to an OD₆₀₀ of approximately 0.5, and mixed in equal volumes. The mixture was spread onto SPA plates and incubated upside down at 30°C for 2–3 days to induce meiosis and sporulation. For the fertility assay, 5–10 μL of propidium iodide (PI, 1 mg/mL) was added to 50 μL of H
2
O, and cells were scraped from the SPA plates and suspended in the PI mix. The mixture was incubated at room temperature for 30 minutes, followed by gentle centrifugation to collect the cells. Fluorescence microscopy was then used for observation and imaging (
Nuckolls et al., 2017
).
Spot assay
The yeast strains were cultured in YPD liquid medium at 30℃ with shaking at 200 rpm. The overnight cultures were transferred to fresh YPD liquid medium and grown to an OD
600
value of ~3. Cells were collected by centrifugation, and the OD
600
was adjusted to 3. Subsequently, the strains were diluted in five 10-fold steps to 10
–5
, and 1.5 μL of each dilution were spotted on the surface of YPD and YPG solid media. The plates were incubated at 30℃, and the growth of colonies was observed.
Data availability
All the data were available in the main text and supplemental information.
References
Experiments on plant hybrids by gregor mendel
Genetics
204
:407–422.
PubMed
Google Scholar
Strength and tempo of selection revealed in viral gene genealogies
BMC Evolutionary Biology
11
:220.
PubMed
Google Scholar
Retrotransposons and their recognition of pol II promoters: a comprehensive survey of the transposable elements from the complete genome sequence of
Schizosaccharomyces pombe
Genome Research
13
:1984–1997.
PubMed
Google Scholar
A suppressor of a wtf poison-antidote meiotic driver acts via mimicry of the driver’s antidote
PLOS Genetics
14
:e1007836.
PubMed
Google Scholar
Genetic villains: killer meiotic drivers
Trends in Genetics
34
:424–433.
PubMed
Google Scholar
Dramatically diverse
Schizosaccharomyces pombe
wtf meiotic drivers all display high gamete-killing efficiency
PLOS Genetics
16
:e1008350.
PubMed
Google Scholar
Atypical meiosis can be adaptive in outcrossed
Schizosaccharomyces pombe
due to wtf meiotic drivers
eLife
9
:e57936.
Google Scholar
Why is mendelian so exact
BioEssays
13
:305–312.
PubMed
Google Scholar
Software
The igraph software package for complex network research
InterJournal. Complex Systems.
Software
Igraph: network analysis and visualization in r
Igraph.
The
wtf
meiotic driver gene family has unexpectedly persisted for over 100 million years
eLife
11
:e81149.
PubMed
Google Scholar
Killer meiotic drive and dynamic evolution of the wtf gene family
Molecular Biology and Evolution
36
:1201–1214.
PubMed
Google Scholar
Centromere-associated meiotic drive and female fitness variation in Mimulus
Evolution; International Journal of Organic Evolution
69
:1208–1218.
PubMed
Google Scholar
New software for the fast estimation of population recombination rates (FastEPRR) in the genomic era
G3: Genes, Genomes, Genetics
6
:1563–1571.
PubMed
Google Scholar
The fitness advantage of commercial wine yeasts in relation to the nitrogen concentration, temperature, and ethanol content under microvinification conditions
Applied and Environmental Microbiology
80
:704–713.
PubMed
Google Scholar
Unifying the epidemiological and evolutionary dynamics of pathogens
Science
303
:327–332.
PubMed
Google Scholar
UFBoot2: improving the ultrafast bootstrap approximation
Molecular Biology and Evolution
35
:518–522.
PubMed
Google Scholar
A large gene family in fission yeast encodes spore killers that subvert Mendel’s law
eLife
6
:e26057.
PubMed
Google Scholar
The role of selfish genetic elements in eukaryotic evolution
Nature Reviews. Genetics
2
:597–606.
PubMed
Google Scholar
Application of phylogenetic networks in evolutionary studies
Molecular Biology and Evolution
23
:254–267.
PubMed
Google Scholar
The evolution of gene duplications: classifying and distinguishing between models
Nature Reviews. Genetics
11
:97–108.
PubMed
Google Scholar
ModelFinder: fast model selection for accurate phylogenetic estimates
Nature Methods
14
:587–589.
PubMed
Google Scholar
MAFFT multiple sequence alignment software version 7: improvements in performance and usability
Molecular Biology and Evolution
30
:772–780.
PubMed
Google Scholar
Improved algorithmic complexity for the 3SEQ recombination detection algorithm
Molecular Biology and Evolution
35
:247–251.
PubMed
Google Scholar
The selfish Segregation Distorter gene complex of
Drosophila melanogaster
Genetic
192
:33–53.
PubMed
Google Scholar
The ecology and evolutionary dynamics of meiotic drive
Trends in Ecology & Evolution
31
:315–326.
PubMed
Google Scholar
Book
The Origins of Genome Architecture
Sunderland (MA): Sinauer Associates.
Google Scholar
Segregation distorters
Annual Review of Genetics
25
:511–557.
PubMed
Google Scholar
IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era
Molecular Biology and Evolution
37
:1530–1534.
PubMed
Google Scholar
Molecular genetic analysis of fission yeast
Schizosaccharomyces pombe
Methods in Enzymology
194
:795–823.
https://doi.org/10.1016/0076-6879(91)94059-l
PubMed
Google Scholar
wtf
genes are prolific dual poison-antidote meiotic drivers
eLife
6
:e26033.
PubMed
Google Scholar
The
wtf4
meiotic driver utilizes controlled protein aggregation to generate selective cell death
eLife
9
:e55694.
PubMed
Google Scholar
S. pombe
wtf drivers use dual transcriptional regulation and selective protein exclusion from spores to cause meiotic drive
PLOS Genetics
18
:e1009847.
PubMed
Google Scholar
Selfish genetic elements and sexual selection: their impact on male fertility
Genetica
132
:295–307.
PubMed
Google Scholar
FastTree 2--approximately maximum-likelihood trees for large alignments
PLOS ONE
5
:e9490.
PubMed
Google Scholar
Meiotic drive as an evolutionary force
The American Naturalist
91
:105–110.
Google Scholar
Detrimental effects of an autosomal selfish genetic element on sperm competitiveness in house mice
Proceedings. Biological Sciences
282
:20150974.
PubMed
Google Scholar
Population genomics of the wild yeast
Saccharomyces paradoxus
: Quantifying the life cycle
PNAS
105
:4957–4962.
PubMed
Google Scholar
Reactivation of transposable elements following hybridization in fission yeast
Genome Research
32
:324–336.
PubMed
Google Scholar
Fertility costs of meiotic drivers
Current Biology
29
:R512–R520.
PubMed
Google Scholar
PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies
Molecular Ecology Resources
20
:348–355.
PubMed
Google Scholar
Ubiquitination-mediated Golgi-to-endosome sorting determines the toxin-antidote duality of fission yeast wtf meiotic drivers
Nature Communications
14
:8334.
PubMed
Google Scholar
BioAider: An efficient tool for viral genome analysis and its application in tracing SARS-CoV-2 transmission
Sustainable Cities and Society
63
:102466.
PubMed
Google Scholar
Article and author information
Author details
Yan Wang

"This ORCID iD identifies the author of this article:"
0009-0002-5375-6082
Funding
National Natural Science Foundation of China (32270684)
Guan-Zhu Han
National Natural Science Foundation of China (32300511)
Zhen Gong
National Natural Science Foundation of China (32470652)
Guan-Zhu Han
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (32270684 and 32470652 to G-ZH and 32300511 to ZG).
Version history
Sent for peer review:
Preprint posted
:
Reviewed Preprint version 1
:
Reviewed Preprint version 2
:
Version of Record published
:
Cite all versions
You can cite all versions using the DOI
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2024, Wang et al.
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
372
views
14
downloads
0
citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads
(link to download the article as PDF)
Article PDF
Figures PDF
Open citations
(links to open the citations from this article in various online reference manager services)
Mendeley
Cite this article
(links to download the citations from this article in formats compatible with various reference manager tools)
Recombination shapes the diversification of the
wtf
meiotic drivers
eLife
13
:RP100638.





