 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库ATP-driven conformational dynamics reveal hidden intermediates in a heterodimeric ABC transporter
Abstract
ATP-binding cassette (ABC) transporters are essential molecular machines whose conformational dynamics have largely been inferred from ensemble-averaged measurements. Resolving dynamic heterogeneity and transient intermediates, however, requires single-molecule approaches. Here, we use single-molecule Förster resonance energy transfer (smFRET) to directly monitor conformational changes of the heterodimeric type IV ABC transporter TmrAB, a functional homolog of the human antigen transporter TAP. Fluorophores positioned at the nucleotide-binding domains and the periplasmic gate were validated by accessible-volume simulations, fluorescence lifetimes, and ensemble FRET, demonstrating that these reporters reliably track conformational transitions. Single-molecule analysis distinguishes ATP-free and ATP-bound states and reveals ATP-dependent population shifts from nucleotide-free to physiological ATP concentrations. Probing conformational dwell-times further uncovers an unexpectedly long ATP-bound dwell time of ∼300 ms. Using complementary stabilization strategies–including a slow-turnover variant, Mg²⁺ depletion, or substrate trans-inhibition–we resolve a previously hidden outward-facing open state that rapidly interconverts with occluded intermediates under turnover conditions. These results provide the first single-molecule characterization of TmrAB and establish a general framework for dissecting ATP-coupled conformational dynamics in heterodimeric ABC transporters.
Introduction
ATP-binding cassette (ABC) transporters constitute the largest family of primary active membrane transport systems, conserved across all domains of life
1
–
3
. Despite considerable structural diversity, all ABC transporters share a modular architecture comprising two conserved nucleotide-binding domains (NBDs)–the defining hallmark of the family–and two transmembrane domains (TMDs) that form the substrate translocation pathway
3
,
4
. Based on their transmembrane-domain architecture, ABC transporters are classified into seven types that encompass importers, exporters, extractors, and mechanotransmitters
5
. Substrate translocation is driven by large conformational changes that are chemo-mechanically coupled to ATP binding, hydrolysis, and phosphate/ADP release
2
,
3
. ABC transporters play central roles in cellular homeostasis, nutrient uptake, waste removal, and toxin defense. Their dysfunction and misregulation are linked to numerous diseases and drug resistance
6
.
The heterodimeric type IV ABC transporter TmrAB from
Thermus thermophilus
has emerged as a powerful model system due to its exceptional thermal stability and functional homology to the transporter associated with antigen processing (TAP1/2), a key component of adaptive immunity
7
–
9
. Notably, TmrAB shares overlapping peptide specificity with TAP and can restore antigen presentation in TAP-deficient human cells
10
. Its inherent asymmetry, with one catalytically active (canonical) and one inactive (noncanonical) nucleotide-binding site (NBS), provides a unique opportunity to investigate functional specialization and asymmetry in ABC transport mechanisms.
Extensive structural studies, particularly using cryogenic electron microscopy (cryo-EM), have delineated the conformational landscape of TmrAB and yielded a detailed model of its translocation cycle
11
,
12
. In this model, TmrAB fluctuates between inward-facing wide and narrow conformations (IF
wide
and IF
narrow
), characterized by a sealed periplasmic gate (PG) and well-separated NBDs, thereby permitting substrate access to the central binding cavity. ATP binding to both NBDs induces NBD dimerization and drives the transition into the outward-facing states, including an OF open (OF
open
) conformation with an open PG that enables substrate release into the periplasm, as well as an OF occluded (OF
occluded
) state characterized by a sealed PG and dimerized NBDs. Subsequent ATP hydrolysis and phosphate release lead to asymmetric unlocked return states (UR
asym
and UR
asym
*), before the transporter returns to the IF conformation. These UR sates feature as sealed PG, a partially open ADP-bound canonical NBS, and a tightly ATP-occluded noncanonical NBS
11
.
Single-turnover experiments established that ATP binding, rather than hydrolysis, drives the IF-to-OF transition, while phosphate release precedes the OF-to-IF switch
12
,
13
. Complementary ensemble approaches, including pulsed electron–electron double resonance (PELDOR/DEER) spectroscopy, have further characterized ATP-dependent conformational changes
14
,
15
. However, ensemble averaging inherently masks molecular heterogeneity, obscures inactive or misfolded subpopulations, and limits access to kinetic information.
Single-molecule techniques overcome these limitations by resolving conformational dynamics at the level of individual molecules
16
,
17
. In particular, single-molecule Förster resonance energy transfer (smFRET) enables real-time monitoring of protein conformational changes with nanometer precision
18
–
21
. Applied to ABC transporters, smFRET provides a unique opportunity to dissect transport cycles, resolve transient intermediates, and extract kinetic and mechanistic insights that remain inaccessible to ensemble-based measurement approaches
22
–
24
.
Here, we apply total internal reflection fluorescence (TIRF) microscopy combined with alternating laser excitation (ALEX)-based smFRET to detergent-solubilized heterodimeric ABC transporter TmrAB, providing the first single-molecule characterization of this system. By strategically positioning fluorophore pairs, we directly monitor ATP-dependent NBD dimerization and periplasmic gate (PG) opening, quantify conformational state occupancies across ATP concentrations ranging from nucleotide-free to physiological levels (3 mM), and uncover conformational dynamics previously masked by ensemble averaging. Using three orthogonal trapping strategies–(i) a slow-turnover catalytic mutant
11
,
12
, (ii) Mg²⁺ depletion
14
,
25
, and (iii) substrate trans-inhibition
26
,
27
–we resolved a previously hidden outward-facing open (OF
open
) state that rapidly exchanges with the outward-facing occluded (OF
occluded
) state. Distance measurements derived from smFRET closely matched predictions from accessible-volume (AV) simulations, cryo-EM structures, and PELDOR/DEER spectroscopy, confirming that detergent-solubilized TmrAB retains a native-like conformational landscape. Together, these results provide the first single-molecule quantification of conformational state occupancies for a heterodimeric type IV ABC transporter and establish TmrAB as a versatile model for single-molecule studies of ABC transport systems.
Results
Design of FRET-labeled TmrAB variants to probe conformational dynamics
To monitor conformational changes in distinct regions of TmrAB, we engineered FRET variants targeting the nucleotide-binding domains (NBDs) and the periplasmic gate (PG). The NBDs undergo ATP-dependent dimerization followed by post-hydrolysis dissociation, whereas the PG opening and closing controls substrate release into the periplasm
2
,
3
,
11
. Probing both regions provides complementary readouts of cytosolic and periplasmic coupling during the transport cycle.
Labeling positions were selected based on prior PELDOR/DEER studies
15
. The NBD reporter variant (TmrA
C416
B
L458C
, referred to as TmrAB
NBD
) monitors conformational changes at the noncanonical nucleotide-binding site (NBS), while the PG reporter (TmrA
C416A, T61C
B
R56C
, hereafter TmrAB
PG
) tracks PG opening. In TmrAB
NBD
, the native single cysteine (C416) was retained for labeling, whereas in TmrAB
PG
it was substituted by alanine to prevent off-target labeling. Selecting the noncanonical rather than the canonical NBS prevents distinguishing outward-facing occluded (OF
occluded
) from asymmetric unlocked return states (UR
asym
and UR
asym
*)
11
, but reduces the number of resolvable FRET states and thereby simplifies data interpretation.
Both variants were labeled with photostable fluorophores, LD555 (donor) and LD655 (acceptor), containing a 1,3,5,7-cyclooctatetraene moiety to suppress photobleaching and blinking
28
,
29
. Accessible-volume (AV) simulations
30
performed on nine cryo-EM structures
11
confirmed that donor-acceptor distances (
R
DA
) and simulated FRET efficiencies (
E
sim
) fall within the FRET-sensitive range (
Fig. 1
). For TmrAB
NBD
,
E
sim
ranged from 0.62 ± 0.02 (57.9 ± 0.7 Å, NBDs separated) to 0.84 ± 0.01 (45.2 ± 0.4 Å, NBDs dimerized). For TmrAB
PG
,
E
sim
shifted from 0.96 ± 0.02 (30.6 ± 0.4 Å, PG closed) to 0.69 ± 0.03 (53.8 ± 2.0 Å, PG open). These transitions correspond to Δ
E
sim
values of 0.22 and 0.27 and Δ
R
DA
of 12.7 Å and 23.2 Å, for TmrAB
NBD
and TmrAB
PG
, respectively, predicting robust and experimentally resolvable FRET changes.
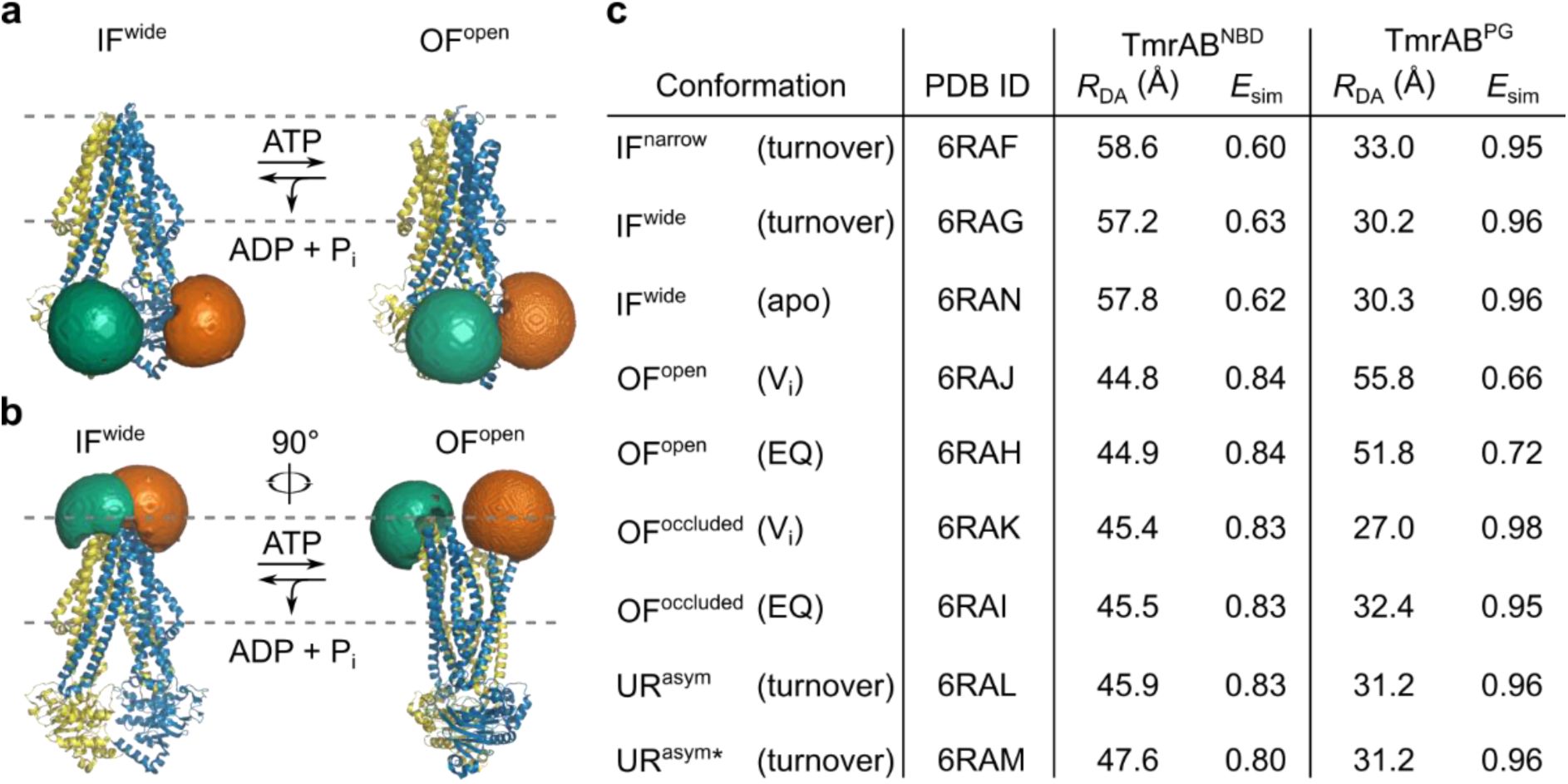
Accessible-volume (AV) simulations of LD fluorophores on TmrAB variants.
AV simulations were performed for LD555 (donor) and LD655 (acceptor) fluorophores attached to the selected TmrAB labeling sites to assess whether donor-acceptor distances are suitable for smFRET measurements
30
.
a
, TmrAB
NBD
(TmrA
C416
B
L458C
) and
b
, TmrAB
PG
(TmrA
C416A, T61C
B
R56C
) in the inward-facing wide (IF
wide
; PDB: 6RAN, left) and outward-facing open (OF
open
; PDB: 6RAH, right) conformations. Approximate membrane position is indicated by the dashed grey line. For all simulations, TmrA is shown in blue with LD655 (orange) and TmrB in yellow with LD555 (green).
c
, AV simulations confirmed that donor-acceptor distances (
R
DA
) remain within the FRET-sensitive range in both conformations, predicting measurable shifts in simulated FRET efficiencies (
E
sim
). Cryo-EM structures of TmrAB reconstituted in lipid nanodiscs
11
were used as templates. Structures were determined either in apo state (apo) or in presence of 3 mM ATP (turnover). Outward-facing open (OF
open
) and outward-facing occluded (OF
occluded
) structures were obtained via orthovanadate trapping (V
i
) or by using the slow-turnover catalytic mutant TmrA
E523Q
B (EQ).

Quality of TmrAB purification and fluorophore labeling.
a
, SDS-PAGE analysis (10%, reducing conditions, Coomassie staining) of successive purification steps: M, molecular weight marker; L, cell lysate; B, Ni-NTA beads after incubation with lysate; FT, flow-through; W, wash; E, eluted TmrAB; BE, TmrAB after buffer exchange; C, concentrated TmrAB; CFT, concentrator flow-through; 1 and 2, first and second peaks eluted from size-exclusion chromatography (SEC). Only the second peak was used for subsequent FRET experiments.
b
, SEC (Superdex 200 increase 10/300 GL) confirming monodispersity of labeled TmrAB and efficient removal of free fluorophores. Representative chromatogram is shown for TmrAB
PG
.
c
, ATP hydrolysis activity of purified wild-type TmrAB (60 nM TmrAB
wt
) measured at 40 °C for 7 min. Released inorganic phosphate (P
i
) was quantified using the Malachite Green assay. Data were fitted to a Michaelis-Menten model, yielding
K
m
= 0.97 ± 0.28 mM and
k
cat
= 2.57 ± 0.38 s
−1
.
d
–
f
, Analytical SEC (Superdex 200 increase 3.2/300) used to determine fluorophore labeling efficiencies for each variant: (
d
) TmrAB
NBD
, LD555: ∼55%, LD655: ∼53%; (
e
) TmrAB
PG
, LD555: ∼43%, LD655: ∼52%; and (
f
) TmrAB
PG_EQ
, LD555: ∼42%, LD655: ∼52%.

FRET capabilities of labeled TmrAB variants.
a
, Time-correlated single-photon counting histograms of LD555 (left) and LD655 (middle) measured under three conditions: free dye in buffer (black), LD555/LD655-labeled TmrAB
NBD
(orange), and LD555/LD655-labeled TmrAB
PG
(blue). Amplitude-weighted average fluorescence lifetimes are summarized in the table (right), confirming sufficient rotational freedom for reliable FRET measurements.
b–d
, Ensemble donor-exited emission spectra (550–700 nm, excitation 520 nm) of (
b
) TmrAB
NBD
, (
c
) TmrAB
PG
, and (
d
) the slow-turnover variant TmrAB
PG_EQ
, stochastically labeled with LD555/LD655 and incubated with increasing ATP concentrations. Spectra are normalized to donor intensity in the apo state. ATP-dependent donor quenching and acceptor sensitization indicate that all variants retain FRET capability.
e–g
, Fractional fluorescence changes, (
F
-
F
₀)/
F
₀, where
F
is acceptor emission intensity and
F
₀ is the intensity in the apo state, plotted as a function of ATP concentration for (
e
) TmrAB
NBD
, (
f
) TmrAB
PG
, and (
g
) TmrAB
PG_EQ
. Data were fitted with a hyperbolic binding model to determine apparent
K
d,
ATP
values, consistent with ensemble FRET measurements of ATP binding.
Additionally, we employed a slow-turnover TmrAB variant that reports on PG opening (TmrA
C416A, E523Q, T61C
B
R56C
, hereafter TmrAB
PG_EQ
). Substituting the catalytic glutamate with glutamine removed the carboxylate required to activate water for nucleophilic attack on ATP, thus drastically reducing the rate of ATP hydrolysis. This mutation slows down the catalytic turnover (∼1000-fold) to a half-life of approximately 25 min at 45 °C
11
,
12
,
14
,
25
, enabling stabilization of ATP-bound outward-facing conformations.
TmrAB variants are suitable for FRET studies
TmrAB variants were expressed in
E. coli
and purified using immobilized metal-affinity chromatography. SDS-PAGE and size-exclusion chromatography (SEC) confirmed high sample purity and monodispersity (
Fig. 1–Fig. S1a,b
). ATPase assays of TmrAB
wt
verified that enzymatic activity was fully retained after purification, yielding a Michaelis-Menten constant (
K
m
) of 0.97 ± 0.28 mM and catalytic ATP turnover rate (
k
cat
) of 2.57 ± 0.38 s
−1
at 40 °C (
Fig. 1–Fig. S1c
).
Cysteine-maleimide labeling of detergent-solubilized TmrAB variants achieved site-specific labeling efficiencies exceeding 90% (
Fig. 1–Fig. S1d–f
). Fluorescence lifetime (
τ
) analysis of conjugated fluorophores confirmed that their photophysical properties were preserved and that they retained sufficient rotational freedom for reliable FRET measurements.
τ
histograms of both conjugated and free fluorophores were fitted with a biexponential decay model, from which amplitude-weighted average lifetimes were calculated. For TmrAB
NBD
, average
τ
values were 0.93 ± 0.02 ns (LD555) and 1.52 ± 0.01 ns (LD655), while TmrAB
PG
exhibited average
τ
values of 0.95 ± 0.02 ns (LD555) and 1.65 ± 0.01 ns (LD655) (
Fig. 1–Fig. S2a
). By comparison, free dyes in buffer displayed lifetimes of 1.11 ± 0.02 ns (LD555) and 1.29 ±0.01 ns (LD655). Because the fluorescence lifetimes of both, the conjugated dyes and the free dyes, remain on the ∼1 ns timescale, we conclude that the fluorophores remain photophysically active and are not affected by protein-induced quenching
17
,
31
. Moreover, the measured lifetimes on the nanosecond timescale are only marginally affected and, most importantly, identical between the TmrAB variants, indicating dynamics orientational averaging of the transition dipoles and confirming that the labeled constructs are suitable for quantitative FRET studies
32
–
34
.
Ensemble ATP titration (0–10 mM ATP) revealed the expected concentration-dependent donor quenching and acceptor sensitization (
Fig. 1–Fig. S2b–d
). ATP-induced fractional fluorescence changes provided as a quantitative readout of conformational transitions, allowing estimation of equilibrium dissociation constants for ATP binding (
K
d,
ATP
) to labeled TmrAB variants (
Fig. 1–Fig. S2e–g
). The measured apparent
K
d,
ATP
values––51 ± 38 μM for TmrAB
NBD
, 68 ± 25 μM for TmrAB
PG
, and 95 ± 26 μM for the slow-turnover variant TmrAB
PG_EQ
––are in good agreement with previously reported values (∼100 µM for TmrA
E523Q
B)
12
, indicating that fluorophore labeling does not perturb ATP binding. Notably, TmrAB
PG_EQ
exhibited a larger shift in ATP-induced fluorescence change than TmrAB
PG
, consistent with stabilization of the ATP-bound conformation and reduced catalytic turnover.
ATP-induced conformational switching resolved by single-molecule FRET
TmrAB variants were site-specifically immobilized on PEGylated coverslips using a conformation-independent, TmrB-specific nanobody (Nb9F10
S63C
)
11
conjugated to maleimide-PEG
11
-biotin (
Fig. 2a
). Previous studies confirmed that this nanobody does not perturb TmrAB transport or ATPase activity
11
,
25
. Donor and acceptor photons were recorded by a total-internal reflection fluorescence (TIRF) microscope using alternating laser excitation (ALEX; NanoImager) at 40 °C (
Fig. 2b
). Single-molecule localization, fluorescence-trajectory extraction, and background correction were performed using NanoImager software, followed by DeepFRET-based machine-learning trace classification and corrections for donor leakage, direct acceptor excitation, and difference in detection-efficiency
17
,
35
,
36
(
Fig. 2–Fig. S1
and
2
)
. FRET efficiency (
E
) and stoichiometry (
S
) were calculated from the corrected fluorescence-trajectories (see Methods,
Eq. 1
and
Eq. 2
).
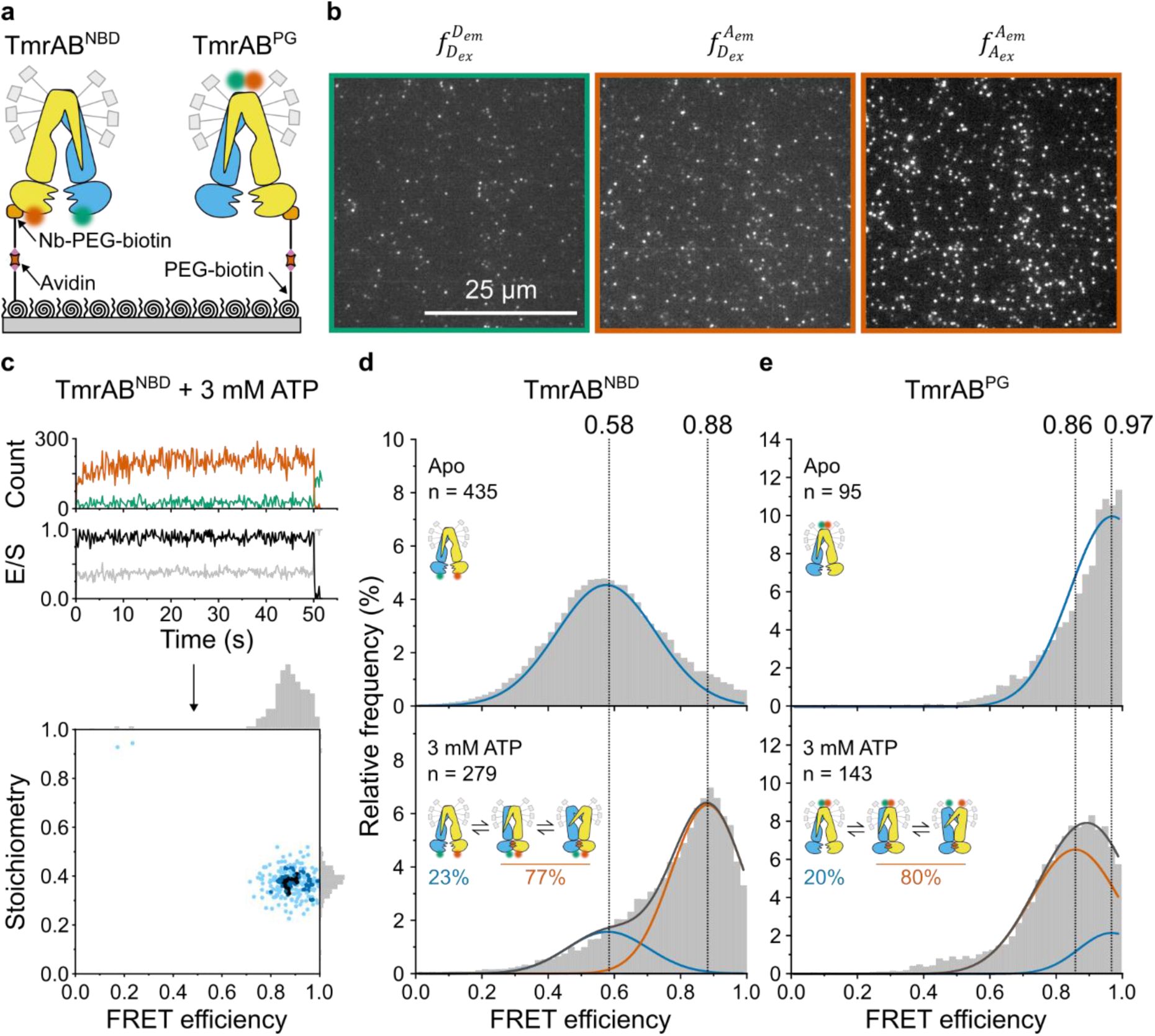
ATP-induced conformational changes of TmrAB analyzed by smFRET.
a
, Experimental setup. TmrAB
NBD
(left) and TmrAB
PG
(right) variants were labeled with LD555/LD655 and tethered to PEGylated coverslips via a biotinylated anti-TmrB nanobody (Nb9F10
S63C
).
b
, smFRET imaging. Samples were recorded using total internal reflection fluorescence (TIRF) microscopy with alternating laser excitation (ALEX; donor: 532 nm; acceptor: 640 nm). Emission was collected in two channels (donor: 498-620 nm; acceptor: 662-710 nm). Donor fluorescence (
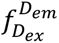
donor excitation at donor emission), sensitized acceptor fluorescence (
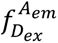
donor excitation at acceptor emission), and direct acceptor fluorescence (
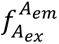
acceptor excitation at acceptor emission) were used to calculate FRET efficiency (
E
) and stoichiometry (
S
). Fluorescence time traces were analyzed with DeepFRET
35
.
c
, Representative TmrAB
NBD
trace in ATP-bound state (top) and corresponding FRET efficiency/stoichiometry plot (bottom). Donor emission upon donor excitation is shown in green as the number of detected photons per frame (counts), acceptor intensity upon donor excitation in orange, FRET efficiency (
E
) in black, and stoichiometry (
S
) in grey. The slight increase in donor fluorescence observed during the first seconds of acquisition reflects photophysical equilibration of the fluorophore and instrumental stabilization and does not affect FRET efficiency or stoichiometry, which are ratio-based and remain constant over time
17
.
d
,
e
, Population analysis. FRET efficiency (
E
) histograms for (
d
) TmrAB
NBD
and (
e
) TmrAB
PG
are shown for the apo state (top) and ATP-bound state (bottom; 3 mM ATP). Histograms were fitted with two Gaussian populations corresponding to the apo state (blue; defined from apo measurements) and the ATP-bound state (orange; defined from two-component fits at saturating ATP). Dotted vertical lines indicate mean
E
values of each population. Population fractions, calculated from Gaussian areas, are summarized schematically in each panel.
Representative smFRET traces of TmrAB
NBD
.
Representative single-molecule FRET (smFRET) traces of TmrAB
NBD
were recorded (
a
) in the ATP-free state and (
b, c
) in the presence of 3 mM ATP. Hidden Markow modeling (HMM) was applied to classify traces into (
b
) static and (
c
) dynamic, based on the absence or presence of transitions between ATP-free and ATP-bound conformational states. Donor fluorescence intensity upon donor excitation is shown in green, acceptor fluorescence intensity upon donor excitation in orange, FRET efficiency (
E
) in black, and stoichiometry (
S
) in grey.
Representative smFRET traces of TmrAB
PG
.
Representative single-molecule FRET (smFRET) traces of TmrAB
PG
were recorded (
a
) in the ATP-free state and (
b, c
) in the presence of 3 mM ATP. Hidden Markow modeling (HMM) was applied to classify traces into (
b
) static and (
c
) dynamic, based on the absence or presence of transitions between ATP-free and ATP-bound conformational states. Donor fluorescence intensity upon donor excitation is shown in green, acceptor fluorescence intensity upon donor excitation in orange, FRET efficiency (
E
) in black, and stoichiometry (
S
) in grey.
FRET efficiency (
E
) histograms revealed two Gaussian populations corresponding to the apo and ATP-bound states (
Fig. 2d,e
). In the absence of ATP, only the apo population was observed, whereas addition of 3 mM ATP induced the appearance of a second ATP-bound population. For TmrAB
NBD
, the apo and ATP-bound populations exhibited mean
E
values of 0.58 and 0.88 (Δ
E
= 0.30), respectively, with ∼77% of molecules occupying the ATP-bound state. For TmrAB
PG
, mean
E
values were 0.97 (apo) and 0.86 (ATP-bound) (Δ
E
= 0.11), with ∼80% of molecules in the ATP-bound state.
Distance estimates calculated using a Förster radius of
R
0
= 63.5 Å (ref.
37
) yielded apparent distances of 60.2 Å (apo) and 45.6 Å (ATP-bound) for TmrAB
NBD
, and 35.6 Å (apo) and 46.9 Å (ATP-bound) for TmrAB
PG
. For TmrAB
NBD
, the experimentally derived Δ
R
of 14.6 Å closely agreed with the AV simulations. In contrast, the smaller Δ
R
of 11.4 Å observed for TmrAB
PG
deviated from simulated values, indicating that the ATP-bound population at this site represents a mixture of rapidly interconverting conformations rather than a single well-defined state.
To assess the ATP sensitivity, we quantified conformational responses across a wide range of ATP concentrations, spanning well below the reported
K
d,
ATP
(∼100 µM for TmrA
E523Q
B)
12
up to physiologically relevant levels (3 mM ATP). smFRET measurements revealed dose-dependent population shifts: TmrAB
NBD
transitioned from a low-FRET apo state (
E
= 0.58) to high-FRET ATP-bound state (
E
= 0.88), whereas TmrAB
PG
shifted from a high-FRET apo state (
E
= 0.97) to a lower-FRET ATP-bound state (
E
= 0.86) (
Fig. 3a,c
). Langmuir isotherm fits yielded
K
d,
ATP
values of 13 ± 1 μM for TmrAB
NBD
and 2 ± 1 μM for TmrAB
PG
(
Fig. 3b,d
), indicating saturation at ATP concentrations well below physiological levels (3 mM). For the TmrAB
PG
variant, population quantification and subsequent determination of
K
d,
ATP
are unreliable due to insufficient separation of the two FRET populations below 1 mM ATP, consistent with the smaller Δ
E
observed for this labeling configuration.

ATP-dependent shifts in smFRET populations of TmrAB.
a
,
c
, Increasing ATP concentrations gradually redistributed the population between apo and ATP-bound conformations for (
a
) TmrAB
NBD
and (
c
) TmrAB
PG
. FRET efficiency (
E
) histograms were fitted with two Gaussian populations corresponding to the ATP-free state (blue: defined from apo samples) and the ATP-bound state (orange; determined from fits at saturating ATP). Dotted vertical lines indicate the mean
E
values of each population. Relative proportion fractions, calculated from Gaussian areas, are summarized schematically in each panel.
b
,
d
, ATP-binding curves were obtained by plotting the fraction of molecules in the ATP-bound states as a function of ATP concentration for (
b
) TmrAB
NBD
(reporting NBD dimerization) and (
d
) TmrAB
PG
(reporting PG opening). Data were fitted with a Langmuir isotherm to determine the apparent dissociation constant
K
d,
ATP
of each variant.
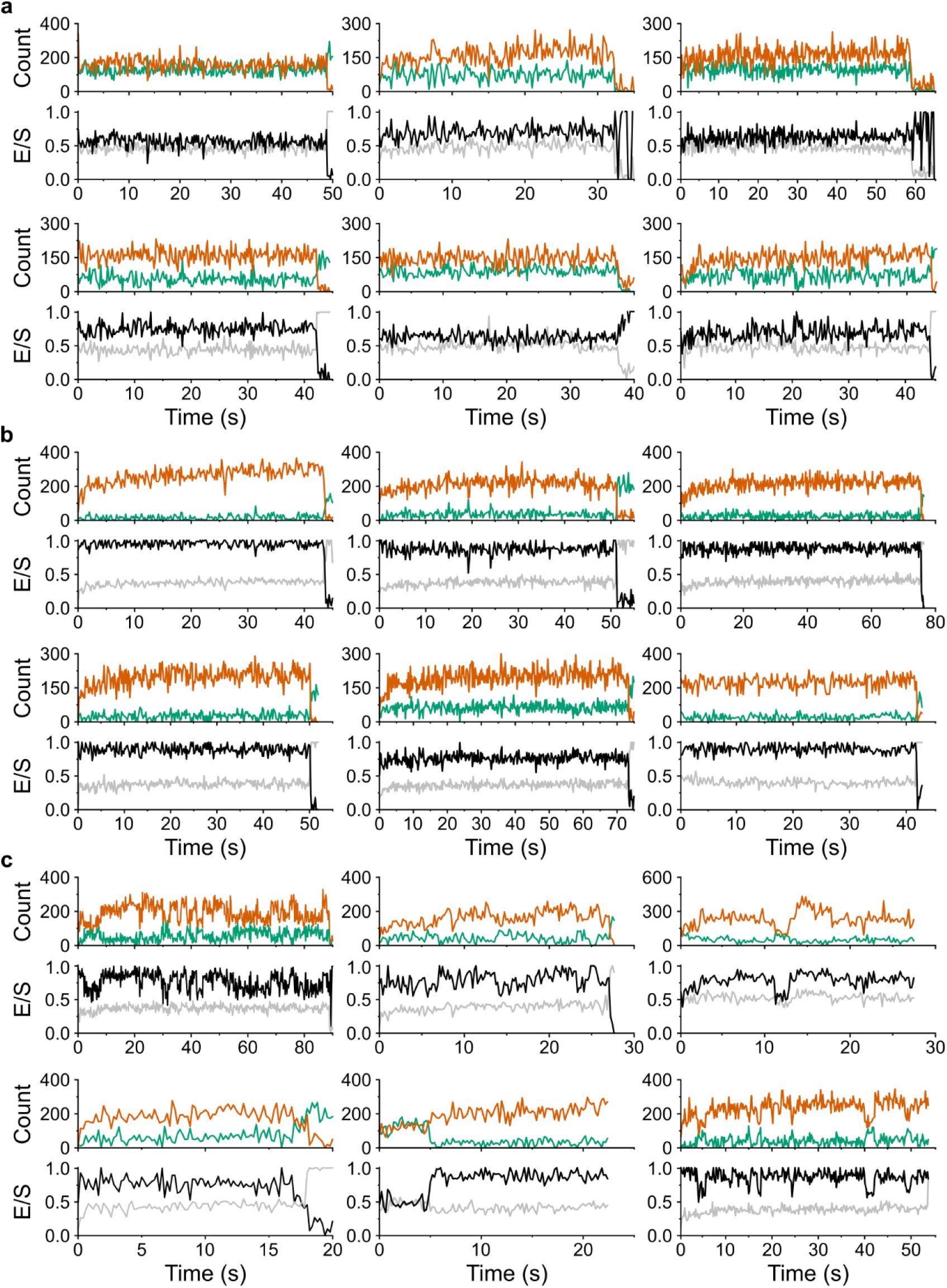
Trapping of TmrAB
PG
reveals a previously hidden outward-facing open state
At physiological ATP concentrations (3 mM), ∼80% of TmrAB
PG
molecules populated the ATP-bound state (
E
= 0.86), closely matching the ATP-bound population (∼77% of NBD-dimerized) observed for TmrAB
NBD
(
Fig. 3
). This agreement indicates that both labeling strategies consistently report ATP-dependent conformational changes. However, the ATP-induced shift observed for the periplasmic-gate reporter TmrAB
PG
(Δ
E
= 0.11, Δ
R
= 11.4 Å;
Fig. 3c
) was substantially smaller than predicted by AV simulations (Δ
E
= 0.27, Δ
R
= 23.2 Å;
Fig. 1c
). This discrepancy indicates that the ATP-bound population at
E
= 0.86 represents an unresolved ensemble, potentially comprising OF
open
and OF
occluded
conformations, as well as the post-hydrolysis asymmetric unlocked return states (UR
asym
and UR
asym
*), which are clearly indistinguishable from OF
occluded
within the current FRET geometry
11
.
To test whether these states are kinetically unresolved by smFRET, we applied three complementary strategies to arrest the OF
open
conformation of TmrAB
PG
: (i) a slow-turnover catalytic mutant (TmrAB
PG_EQ
), (ii) Mg²⁺ depletion using EDTA, and (iii) reverse inhibition by high concentrations of the substrate peptide RRYQKSTEL (R9L) (
Fig. 4
). Slow-turnover variants have previously enabled structural separation of OF
open
and OF
occluded
states
11
,
12
,
14
,
25
. Mg²⁺ depletion blocks ATP hydrolysis while preserving ATP binding, allowing rapid and reversible trapping
25
. We further hypothesized that trans-inhibition by peptide binding sterically restricts PG closure and is therefore expected to stabilize OF
open
in a dose-dependent manner
26
,
27
.

Identification of the outward-facing open (OF
open
) conformation.
a–c
, Three complementary approaches were employed to resolve OF
open
state: (
a
) the slow-turnover variant TmrAB
PG_EQ
, (
b
) imaging in Mg
2+
-free buffer supplemented with EDTA, and (
c
) stabilizing via reverse inhibition using high concentrations of peptide substrate. FRET efficiency (
E
) histograms were fitted with three Gaussian populations corresponding to the ATP-free state (blue), the ATP-bound state (orange), and the stabilized OF
open
state (green). All three strategies revealed a distinct OF
open
population. Dotted vertical lines indicate the mean
E
values of each population. Relative proportion fractions, calculated from Gaussian areas, are summarized schematically in each panel.
d
, Comparison of inter-residues distances. Distances (Å) between selected residues on the NBDs and PG of TmrAB were determined by multiple methods: smFRET values (this study) for detergent-solubilized TmrAB; cryo-EM distances (C
β
–C
β
) from nanodisc-reconstituted TmrAB (PDB 6RAH, 6RAN)
11
; accessible-volume (AV) simulation distances for nanodisc-reconstituted TmrAB (this study); and PELDOR/DEER distances from detergent-solubilized TmrAB
15
.
In conditions lacking ATP, either in the absence of nucleotides or in the presence of ADP (3 mM ADP), the slow-turnover variant TmrAB
PG_EQ
populated a single high-FRET state (
E
= 0.97) (
Fig. 4a, top and middle
). These results indicate that ADP binding alone is insufficient to promote NBD dimerization nor PG opening, consistent with previous biochemical and structural observations
11
,
12
. Upon ATP addition (3 mM ATP), however, the conformational landscape diverged sharply from that of wild-type TmrAB
PG
. Instead of the two-state distribution observed for wild-type (apo:
E
= 0.97, ∼20%; ATP-bound:
E
= 0.86, ∼80%) (
Fig. 2e
, bottom), TmrAB
PG_EQ
exhibited three well-resolved populations with mean
E
values of
E
= 0.97 (∼14%),
E
= 0.86 (∼55%), and
E
= 0.63 (∼31%) (
Fig. 4a
, bottom).
As in wild-type TmrAB
PG
, the high-FRET population (
E
= 0.97) corresponds to the IF state, whereas the intermediate-FRET population (
E
= 0.86) likely represents a dynamic equilibrium of OF
open
and OF
occluded
conformations, as suggested by cryo-EM analyses
11
. Post-hydrolysis return states (UR
asym
and UR
asym
*) are expected to be minimally populated in TmrAB
PG_EQ
due to its drastically reduced ATP hydrolysis rate
12
. Notably, the low-FRET ATP-bound population (
E
= 0.63) was entirely absent in wild-type. The transition from
E
= 0.97 to
E
= 0.63 corresponds to Δ
E
= 0.34 and Δ
R
= 22.5 Å, in close agreement with the IF◊OF
open
distance predicted by AV simulations (Δ
R
= 23.2 Å;
Fig. 1
), thereby postulating
E
= 0.63 as the OF
open
conformation.
Mg²⁺ depletion independently reproduced this three-state landscape. In the absence of Mg
2+
, wild-type TmrAB
PG
transitioned from a single apo population (without ATP;
E
= 0.97) (
Fig. 4b
, top) to three ATP-bound populations (3 mM ATP;
E
= 0.97, ∼13%;
E
= 0.86, ∼56%;
E
= 0.63, ∼31%) (
Fig. 4b
, middle) closely resembling those observed for the slow-turnover variant (
Fig. 4a
, bottom). Reintroducing Mg²⁺ abolished the
E
= 0.63 population and restored the wild-type two-state distribution (
Fig. 4b
, bottom). This reversibility confirmed that the ATP-bound OF
open
state (
E
= 0.63) is selectively revealed only when ATP hydrolysis is prevented.
Finally, we tested whether periplasmic substrate binding shifts the conformational equilibrium of wild-type TmrAB
PG
toward OF
open
. In the presence of ATP (3 mM), increasing concentrations of peptide substrate (0.3–2 mM R9L) progressively enriched the
E
= 0.63 population from ∼20% to ∼38% (
Fig. 4c
). This dose-dependent stabilization mirrors trans-inhibition behavior reported for human TAP1/2 and reflects the upper substrate-loading capacity of the transporter
26
.
Distance changes derived from smFRET closely match AV simulations, cryo-EM structures (PDB 6RAH, 6RAN)
11
, and DEER/PELDOR measurements
14
(
Fig. 4d
), together validating assignment of the
E
= 0.63 population as the OF
open
conformation.
Kinetics and thermodynamics of the transport cycle
ALEX-smFRET data were acquired with an effective temporal resolution of 200 ms (100 ms per excitation channel). Shorter integration times compromised the signal-to-noise ratio and precluded reliable FRET determination. To quantify conformational dynamics, we applied Hidden Markov Modeling (HMM) using MASH-FRET
38
, classifying traces as either static (single FRET state) or dynamic (multiple states). Approximately 95% of traces in each condition were classified as static, indicating that most conformational transitions occur at or below our temporal resolution.
Although individual transitions could not be directly resolved, population-based analysis (
Fig. 3
), combined with biochemical turnover measurements (
Fig. 1–Fig. S1c
), allowed estimation of ATP-bound dwell times. At saturating ATP conditions well above the apparent
K
d,
ATP
(3 mM, 40 °C), wild-type TmrAB exhibited a catalytic turnover rate of
k
cat
= 2.57 ± 0.38 s
−1
(
Fig. 1–Fig. S1c
), corresponding to a full transport cycle time (
τ
cycle
) of 395 ± 55 ms. ATP-bound dwell times (
τ
d
) were derived from population ratios obtained from Gaussian fits of the FRET efficiency histograms (
Fig. 3
; see Methods,
Eq. 3
and
Eq. 4
). These analyses yielded ATP-bound dwell times of 304 ± 43 ms for TmrAB
NBD
(∼77% ATP-bound) and 316 ± 44 ms for TmrAB
PG
(∼80% ATP-bound), with the remaining ATP-free intervals (∼20–23%) accounting for 91 ms and 79 ms of the cycle, respectively.
Together, these measurements establish a quantitative, single-molecule description of conformation state occupancies and dwell times throughout the catalytic cycle of a heterodimeric ABC transporter under active turnover conditions, providing valuable insights into the dynamic landscape of its translocation cycle.
Discussion
smFRET has become an indispensable tool for dissecting conformational dynamics of membrane proteins, including receptors, ion channels, and transporters, by directly linking structural transitions to functional states With the ABC transporter family, however, smFRET studies have largely been confined to monomeric or homodimeric systems
22
–
24
, leaving the dynamic behavior of asymmetric, heterodimeric transporter comparatively unexplored. Here, we apply smFRET to the heterodimeric type IV ABC transporter TmrAB, extending single-molecule analysis to an asymmetric transporter system and uncovering dynamic features of the transport cycle that are inaccessible to ensemble-averaged approaches.
By positioning FRET reporters at the nucleotide-binding domains (NBDs) and periplasmic gate (PG), we directly monitored ATP-dependent coupling between chemical energy input and global conformational rearrangements. Importantly, these structural rearrangements are not strictly correlated: NBD dimerization can give rise to either an outward-facing open (OF
open
) or occluded (OF
occluded
) conformation
11
,
12
. This decoupling underscores the need to monitor both cytosolic and periplasmic regions to resolve the transport mechanism.
Labeling sites previously validated for PELDOR/DEER spectroscopy
10
,
14
,
15
were adapted for smFRET and rigorously benchmarked using accessible-volume (AV) simulations
30
, fluorescence lifetime analysis, and ensemble FRET titrations. This additional validation is essential because fluorophores impose stricter steric and rotational constraints than nitroxide spin labels
32
. Collectively, these controls demonstrate that fluorophore attachment preserves native-like conformational behavior and ATP binding, establishing TmrAB as a robust system for quantitative single-molecule analysis. Consistent with this conclusion, fluorescence lifetime analysis showed no evidence for substantial protein-fluorophore quenching or restricted dye motion, as indicated by donor lifetime shortening and prolonged acceptor lifetimes characteristic of efficient energy transfer
17
.
Single-molecule measurements resolved two dominant FRET populations corresponding to apo and ATP-bound states for both reporter variants TmrAB
NBD
and TmrAB
PG
. ATP titrations spanning concentrations well below the reported apparent
K
d,
ATP
(∼100 µM for TmrA
E523Q
B)
12
up to physiologically relevant levels (3 mM ATP) revealed gradual, concentration-dependent shifts between these populations, demonstrating the high sensitivity of the FRET constructs to ATP binding. Notably, apparent
K
d,
ATP
values derived from smFRET (2–13 µM) were substantially lower than those obtained from ensemble FRET measurements (50–100 µM). This difference likely reflects the inherent selectively of single-molecule analyses for properly folded and catalytically competent transporters. In smFRET experiments, aggregated or inactive species can be identified and excluded during trace selection based on fluorescence intensity, stoichiometry, and photobleaching behavior, thereby enriching the analyzed population for functional molecules.
Crucially, the ATP-bound population may reflect a rapidly interconverting ensemble of conformations, rather than a single static structure, potentially including OF
open
, OF
occluded
, and post-hydrolysis unlocked return states (UR
asym
and UR
asym
*). These transitions occur faster than the ∼200 ms temporal resolution of our measurements, resulting in averaged FRET efficiencies under turnover conditions. Using three independent trapping strategies—slow-turnover catalysis, Mg²⁺ depletion, and substrate trans-inhibition—we stabilized and directly resolved a previously hidden OF
open
conformation. The associated distance changes are generally consistent with cryo-EM structures
11
, PELDOR/DEER data
10
,
14
,
15
, and simulation-based predictions, suggesting that detergent-solubilized TmrAB samples a largely native-like conformational landscape.
Although detergent-solubilized and lipid nanodisc-reconstituted TmrAB exhibit similar global conformational states, the conformational space accessible to attached fluorophores may be differentially influenced by membrane-associated environments
19
,
39
,
40
. In particular, fluorophores attached near the periplasmic gate may experience steric restrictions due to partial overlap with the membrane region, as suggested by AV simulations. While such effects are negligible under the detergent conditions used here, they should be carefully evaluated in future studies employing membrane-embedded systems. Single-molecule measurements further revealed that addition of the peptide substrate induces concentration-dependent shifts in the conformational equilibrium. Increasing substrate concentrations progressively stabilized the OF
open
state, consistent with trans-inhibition behavior observed in human TAP1/2
26
and bovine ABCC1
27
, and reflecting the finite substrate-loading capacity of the transporter
26
.
Quantitative deconvolution of FRET populations enabled direct determination of conformational state occupancies under physiological ATP concentrations. During active turnover, TmrAB populates the IF state (∼20%), the OF
open
state (∼25%), and OF
occluded
/post-hydrolysis states (UR
asym
and UR
asym
*) (∼55%) (
Fig. 5
). The current reporter geometries do not allow direct discrimination between OF
occluded
and post-hydrolysis states because fluorophores were placed at the noncanonical nucleotide-binding site. However, contributions from post-hydrolysis states are expected to be minimal for slow-turnover TmrAB variant (TmrAB
PG_EQ
), owing to its drastically reduced ATP hydrolysis rate. To our knowledge, this represents the first single-molecule quantification of conformational equilibria for a heterodimeric ABC transporter under catalytic conditions. While these distributions broadly align with cryo-EM particle classifications
11
,
25
, smFRET resolves fewer IF states than the cryo-EM distinction between IF
wide
and IF
narrow
, and additionally captures ATP-bound intermediates that are challenging to resolve structurally due to rapid interconversion.

Conformational state distribution and catalytic cycle of TmrAB under active turnover.
Schematic of the TmrAB transport cycle summarizing major conformational states and their estimated population distributions under physiological ATP concentrations (3 mM, 40 °C).
a
, The inward-facing apo state (IF
narrow
and IF
wide
; blue arc) accounts for ∼20% of molecules and is characterized by separated NBDs and a cytosol-accessible substrate-binding cavity. Substrate binding stabilizes the IF
wide
conformation
11
. Independent of substrate, ATP binding induces NBD dimerization and transition to the ATP-bound ensemble.
b
,
c
, Under substrate-bound turnover conditions, TmrAB proceeds via the (
b
) OF
occluded
to (
c
) OF
open
state in which the substrate release occurs. Under steady-state turnover, the ATP-bound ensemble rapidly interconverts between substrate-free OF
occluded
and OF
open
accounting for ∼25% of the ATP-bound population (green circle). These transitions occur faster than the ∼200 ms temporal resolution of standard smFRET measurement, resulting in an averaged signal under turnover conditions. OF
occluded
likely serves as an obligate intermediate between IF and OF
open
, preventing substrate backflow by maintaining a substrate-binding cavity occluded during the structural rearrangements of the PG and NBDs. Although a substrate-bound OF
occluded
state has not been directly observed for TmrAB, its existence is supported by structures of the homodimeric type IV transporter BmrA
46
. Reduced ATP hydrolysis or substrate trans-inhibition enables trapping of the transporter in the OF
open.
state.
d
,
e
, ATP hydrolysis and phosphate (P
i
) release generate post-hydrolysis return states: (
d
) UR
asym
and (
e
) UR
asym
*. Subsequent ADP release restores the apo IF conformation, completing the transport cycle. Overall, the ATP-bound phase (
b–e
) represents ∼55% occupancy (orange arc) with an estimated dwell time of ∼310 ms, whereas the apo/ATP-rebinding phase (
a
) lasts ∼90 ms, yielding a total cycle time of ∼400 ms (
k
cat
= 2.57 s⁻¹). TmrA is shown in blue, TmrB in yellow, substrate as a green diamond, and nucleotides as orange symbols. Dotted grey boxes indicate the approximate position of the NBD dimer interface.
All smFRET measurements were performed at 40 °C to maintain consistency with prior biochemical studies and ensure fluorophore stability. At the physiological temperature of
T. thermophilus
(68 °C), absolute rates of ATP turnover and conformational transitions are expected to increase, although relative state occupancies may remain conserved if the underlying free-energy landscape is preserved. Despite a substantial fraction of static single-molecule trajectories, ATP-dependent population shifts and catalytic rates indicate that TmrAB operates near the temporal resolution limit of our measurements. Integrating smFRET-derived state occupancies with biochemical turnover rates yields an ATP-bound dwell time of approximately 300 ms, in good agreement with previous biochemical estimates
9
,
12
.
Emerging microsecond-resolution smFRET approaches offer the potential to directly visualize short-lived intermediates within the transport cycle
41
. Future studies could further benefit from three- or four-color FRET strategies
42
,
43
, which would allow simultaneous monitoring of multiple structural elements. In particular, dual labeling of the NBDs and PG could provide direct detection of the OF
occluded
state, while probes placed at both canonical and noncanonical nucleotide-binding sites could capture post-hydrolysis conformational dynamics.
In summary, this work establishes smFRET as a powerful approach for mapping the dynamic landscape of asymmetric ABC transporters. By quantitatively linking ATP binding, conformational equilibria, and kinetics at the single-molecule level, our study resolved an important aspect of the transport mechanism how chemical energy is transduced into directional transport in heterodimeric ABC systems. Integration of native lipid environments, higher temporal resolution, and substrate engagement will further illuminate the coordination of ATP hydrolysis and substrate translocation during transport.
Methods
Expression, purification, and labeling of TmrAB
His
10
-tagged TmrAB variants were expressed in
E. coli
BL21(DE3) (Thermo Fisher Scientific) as described previously
12
. Cells were grown in high-salt LB media (Carl Roth) supplemented with 100 μg ml
−1
ampicillin (PAA Laboratories) at 37 °C. At an OD
600
of 0.5, expression was induced with 1 mM isopropyl β-D-thiogalactopyranoside (IPTG; Carl Roth), and cultures were incubated for 3 h at 37 °C. Cells were harvested by centrifugation (4,500 ×
g
, 4 °C, 15 min) and stored at –80 °C.
For purification, cell pellets were resuspended in lysis buffer (20 mM HEPES-NaOH pH 7.5, 300 mM NaCl, 50 µg ml
−1
lysozyme, 0.2 mM phenylmethylsulfonyl fluoride (PMSF)) and lysed by sonication. Cell debris was removed by centrifugation (18,000 ×
g
, 4 °C, 35 min), and membranes were collected by ultracentrifugation (100,000 ×
g,
4 °C, 30 min). Membranes were solubilized for 2 h at 4 °C in purification buffer (20 mM HEPES-NaOH pH 7.5, 300 mM NaCl) containing 20 mM n-dodecyl β-D-maltoside (β-DDM; Carl Roth). After ultracentrifugation (100,000 ×
g,
30 min, 4 °C), the supernatant was incubated with Ni-NTA agarose (Bio-Rad) for 1 h at 4 °C. The resin was washed with 20 column volumes of wash buffer (20 mM HEPES-NaOH pH 7.5, 300 mM NaCl, 1 mM β-DDM) containing 50 mM imidazole, and TmrAB was eluted with elution buffer (20 mM HEPES-NaOH pH 7.5, 300 mM NaCl, 1 mM β-DDM, 300 mM imidazole).
For fluorophore labeling, TmrAB variants were conjugated via maleimide chemistry using LD555 and LD655 (Lumidyne Technologies). Labeling was carried out at a 1:10:10 molar ratio of protein to each dye in elution buffer for 3 h at 4 °C. Excess dye was quenched with 2 mM β-mercaptoethanol (Sigma-Aldrich), and the labeled protein was buffer-exchanged into size-exclusion chromatography (SEC) buffer (20 mM HEPES-NaOH pH 7.5, 150 mM NaCl, 1 mM β-DDM) using a Zeba Spin Desalting Column (Thermo Fisher Scientific). Unreacted fluorophores were removed by SEC on a Superdex 200 Increase 10/300 GL column (Cytiva). Labeling efficiency was determined by analytical SEC (Superdex 200 Increase 3.2/300; Cytiva) by monitoring absorbance at 280, 555, and 655 nm. To preserve sample integrity for smFRET measurements, TmrAB was purified and labeled within a single day, stored on ice, and imaged over the following two days.
Time-correlated single-photon counting (TCSPC)
Fluorescence lifetime measurements were performed using a FluoTime 100 spectrometer (PicoQuant) equipped for time-correlated single-photon counting (TCSPC). Experiments were carried out on labeled TmrAB in SEC buffer (20 mM HEPES-NaOH pH 7.5, 150 mM NaCl, 1 mM β-DDM). LD555 and LD655 were excited at 510 nm and 610 nm, respectively. Emission was collected using a 620/60 nm bandpass filter for LD555 and a BG4 700 nm long-pass filter for LD655. Photon arrival times were accumulated until the TCSPC histogram reached a peak count of 50,000 photons. Fluorescence decay curves were analyzed by fitting mono- or bi-exponential decay models using FluoFit software (PicoQuant) and amplitude weighted average of fluorescence lifetime was calculated.
Nanobody production and purification
The nanobody Nb9F10
S63C
was expressed and purified as described previously
11
. Briefly, Nb9F10
S63C
was produced in
E. coli
BL21(DE3) cells grown in Terrific Broth (TB; Carl Roth) supplemented with 100 μg ml
−1
ampicillin at 37 °C. At an OD
600
of 0.6, expression was induced with 1 mM IPTG, followed by overnight incubation at 28 °C. Cells were harvested by centrifugation (4,500 ×
g
, 4 °C, 15 min) and stored at –80 °C. For purification, cell pellets were resuspended in nanobody lysis buffer (25 mM HEPES-NaOH pH 7.4, 300 mM NaCl, 15 mM imidazole, 0.5 mM PMSF) and disrupted by sonication. Cell debris was removed by centrifugation (18,000 ×
g
, 4 °C, 35 min), and the clarified lysate was applied to Ni-NTA agarose equilibrated in potassium phosphate (KP
i
) buffer (25 mM KP
i
pH 6.5, 100 mM KCl, and 0.5 mM tris(2-carboxyethyl) phosphine (TCEP)). Bound nanobody was washed with 10 column volumes (CV) of KP
i
buffer and eluted with 8 CV of elution buffer (25 mM KP
i
pH 6.0, 20 mM KCl, 300 mM imidazole, 0.5 mM TCEP). Eluted fractions were pooled and further purified by cation exchange chromatography (CEX) on a HiTrap SP column (Cytiva) using a linear gradient from low-salt buffer (25 mM KP
i
pH 6.0, 20 mM KCl, 0.5 mM TCEP) to high-salt buffer (25 mM KP
i
pH 6.0, 500 mM KCl, 0.5 mM TCEP). The purified nanobody was concentrated and buffer-exchanged into nanobody SEC buffer (20 mM HEPES-NaOH pH 7.5, 150 mM NaCl) using Zeba spin desalting columns, followed by SEC (Superdex 200 Increase 10/300 GL; Cytiva). For site-specific conjugation, Nb9F10
S63C
was incubated with a biotin-PEG
11
-maleimide linker (Sigma-Aldrich) at a 1.2:1 molar ratio of protein to linker in the presence of 0.5 mM TCEP for 2 h at 4 °C. Excess linker was removed by desalting on Zeba Spin Desalting Columns, followed by a final SEC step (Superdex 200 Increase 10/300 GL).
SDS-PAGE
The purity of TmrAB samples was assessed by SDS-PAGE. Resolving gels (12%) were prepared using 12% (w/v) acrylamide, 0.5 M Tris-HCl (pH 8.8), 0.13% (w/v) SDS, 0.05% (w/v) ammonium persulphate (APS), and 0.25% (v/v)
N,N,N’,N’-
tetramethylethylenediamine (TEMED). Stacking gels contained 4.3% (w/v) acrylamide, 0.5 M Tris/HCl (pH 6.8), 0.09% (w/v) SDS, 0.09% (w/v) APS, and 0.33% (v/v) TEMED. Gels were used immediately or stored at 4 °C for up to 4 weeks. Protein samples were mixed with 4× SDS loading buffer containing dithiothreitol (DTT; Sigma-Aldrich) and heated at 90 °C for 5 min before loading. Electrophoresis was performed at a constant voltage of 120 V using 1× SDS running buffer (25 mM Tris-HCl pH 8.8, 192 mM glycine, 0.1% SDS). Proteins were visualized by staining with InstantBlue
TM
Protein Stain (Abcam) for 1 h at room temperature with gentle agitation and imaged using a Fusion FX system (Vilber).
ATPase activity assay
The ATPase activity of β-DDM-solubilized TmrAB
wt
was quantified using a Malachite Green-based colorimetric assay as described previously
44
. Detergent-solubilized TmrAB (0.6 μM) was incubated in ATPase buffer (20 mM HEPES-NaOH pH 7.5, 150 mM NaCl, 2 mM MgCl
2
, 1 mM β-DDM) containing 3 mM ATP (Sigma-Aldrich) at 40 °C for 7 min. Autohydrolysis controls were prepared by incubation of ATP in ATPase buffer without protein. Reactions were quenched by adding 20 mM H
2
SO
4
, followed by incubation with 3 mM Malachite Green (Thermo Fisher Scientific), 0.2% (v/v) Tween20 (Carl Roth), and 1.5% (w/v) ammonium molybdate (Carl Roth) for 10 min at room temperature. The absorbance at 620 nm was recorded on a CLARIOstar v.5.20 R5 plate reader (BMG LABTECH).
Ensemble FRET measurements
The ATP binding and FRET characteristics of selected TmrAB variants were assessed by ensemble FRET. Labeled TmrAB (100 nM) was incubated with increasing concentrations of ATP at 42 °C for 5 min. Donor-excited emission was recorded from 550–700 nm with an excitation wavelength of 520 nm using a Clariostar v.5.20 R5 plate reader (BMG LABTECH). Acceptor emission intensities at 675 nm were plotted against ATP concentration and fitted with a hyperbolic function to determine the apparent dissociation constant (
K
d,
ATP
) for each variant.
Functionalization of glass slides for single-molecule FRET analysis
Glass coverslips used for TmrAB immobilization in smFRET experiments were functionalized by PEGylation as described previously
45
. Coverslips (Carl Roth) were cleaned by sequential sonication in Milli-Q water and analytical-grade acetone (>99.9%; VWR International), followed by oxygen plasma treatment (0.3 mbar, 80% power, 15 min) using a Zepto plasma cleaner (Diener) and a 10 min incubation in methanol (Avantor, Gliwice, PL). Coverslips were then silanized by incubation for 30 min in a solution of 100 ml methanol, 5 ml acetic acid, and 3 ml 3-aminopropyltrimethoxysilane (APTES; Tokyo Chemical Industry). After silanization, sides were rinsed four times with methanol and dried under a nitrogen stream. Surface functionalization was achieved using a mixture of biotinylated-PEG (4 mol%) and nonbiotinylated-PEG (96 mol%; Rapp Polymer). The PEG solution was sandwiched between two coverslips and incubated overnight in a humidity chamber. Coverslips were then rinsed thoroughly with Milli-Q water and dried under nitrogen. To enhance passivation, a second PEGylation step was performed using 25 mM CH
3
-PEG-NHS (333 Da; Thermo Fisher Scientific) under the same conditions. Finally, slides were rinsed with Milli-Q water, dried under nitrogen, and stored at –20 °C under argon until use.
Single-molecule FRET imaging
Single-molecule FRET (smFRET) experiments were performed using a flow chamber system (Ibidi). Chambers were assembled by placing a biotin-PEG-functionalized glass slide onto a μ-Slide I Luer Family flow channel (Ibidi), with the functionalized surface facing inward. All buffers and Milli-Q water were filtered through 0.2 μm filters (Sigma-Aldrich). Chambers were washed with 1 ml Milli-Q water and incubated with 0.2 mg ml
−1
streptavidin (Sigma-Aldrich) at 4 °C for 30 min to allow binding to the biotin-PEG surface. Unbound streptavidin was removed by washing with 1 ml Milli-Q water. The surface was then treated with 0.3 mg ml
−1
biotinylated-PEG
11
-Nb9F10
S63C
at 4 °C for 45 min, followed flushing with 2 ml SEC buffer (20 mM HEPES-NaOH pH 7.5, 150 mM NaCl, 1 mM β-DDM). Detergent-solubilized, fluorophore-labeled TmrAB (100 nM) was added and incubated at 4 °C for 1 h. Unbound protein was removed by five washes with 1 ml TmrAB-SEC buffer. Chambers were equilibrated with 1 ml of imaging buffer containing 25 mM HEPES-NaOH (pH 7.5), 150 mM NaCl, 3 mM MgCl
2
, 50 mM glucose, 5 mM Trolox, 7.5 U ml
−1
pyranose oxidase, and 1 kU ml
−1
catalase, supplemented with the desired ATP concentration. For EDTA trapping experiments, MgCl
2
was omitted and replaced with 3 mM ethylenediaminetetraacetic acid (EDTA; Sigma-Aldrich). smFRET data were acquired at 40 °C using alternating laser excitation (ALEX) on a total internal reflection fluorescence (TIRF) microscope (NanoImager S, ONI, Oxford, UK). Typically, 600 frames were recorded per region of interest (ROI) with 100 ms exposure time. Laser powers were 0.8 mW cm
−2
(532 nm) and 0.9 mW cm
−2
(640 nm). Data were recorded in 1-min intervals, except for TmrAB
NBD
variant in apo and 3 mM ATP conditions, where 3-min intervals were used to confirm that conformation transitions do not occur on timescales longer that one minute due to the reduced temperature.
Single-molecule FRET data analysis
Single-molecule FRET (smFRET) measurements were performed using alternating laser excitation (ALEX), allowing assignment of detected photons based on both excitation and emission wavelengths. Photon counts for each molecule were extracted using NanoImager software (ONI NanoImager, Development Build) and classified into three detection channels: donor excitation with donor emission (
), donor excitation with acceptor emission (
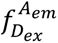
), and acceptor excitation with acceptor emission (
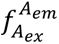
). Traces were analyzed with DeepFRET
35
and manually curated. To minimize bias, a second researcher independently curated traces from both ATP-free and 3 mM ATP samples,


yielding 98% overlap between curations. FRET efficiency (
E
) and stoichiometry (
S
) were calculated as:
Population analysis was performed by constructing one-dimensional histograms of FRET efficiency (
E
) and stoichiometry (
S
) using OriginPro 2024 (OriginLab). Histograms were fitted with two Gaussian distributions corresponding to the ATP-free state (defined from apo samples) and the ATP-bound state (defined by a two-component fit at saturating ATP). Hidden Markov Modeling (HMM) of individual traces was performed using MASH-FRET
38
to distinguish dynamic from static molecules within each sample.
ATP-bound dwell times and distribution of conformational states
The ATP-bound dwell time (
τ
d
) was estimated as:

where
f
ATP-bound
is the fraction of ATP-bound molecules derived from Gaussian fits of the FRET efficiency histograms (
Fig. 3
). This approach assumes (i) ATP hydrolysis occurs exclusively at the canonical NBS, with negligible contribution from the noncanonical site
9
,
11
,
14
, and (ii) the majority of molecules are catalytically competent and continuously cycling.
The distribution of conformational states within ATP-bound FRET population (
E
= 0.86) of TmrAB
PG
under turnover conditions (3 mM ATP) was determined using a two-state model:
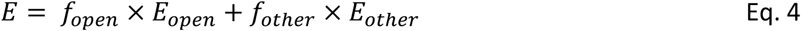
where
f
open
is the fraction of OF
open
state (
E
open
= 0.63) readily resolved under trapping conditions (
Fig. 4
), and
f
other
represents the combined fraction of PG-closed states (OF
occluded
/UR
asym
/UR
asym
*,
E
other
= 0.97).
Data availability
Source data are provided with this paper: DOI:
.
Acknowledgements
This work was supported by the European Research Council (ERC Advanced Grant 101141396 to R.T.), the German Research Foundation via the Collaborative Research Center CRC 1507/P18 to R.T. and the Research Training Group (GRK 1986/B4.7 to R.T.). We thank Jan F.M. Stuke and Jonas Göhmann for their support in automating trace extraction from ONI NanoImager software, Dr. David Glück for guidance on lifetime measurements, and Tobias Nocker for preparing nanobodies used in the study. We are also grateful to the Wachtveitl lab (Goethe University Frankfurt) for access to their FluoTime 100 spectrometer (PicoQuant). Finally, we thank Dr. Rupert Abele, Dr. David Glück, Dr. Simon Trowitzsch, Inga Nold, and Andrea Pott for helpful comments on the manuscript and proofreading.
Additional information
Author contributions
M.P. prepared all TmrAB samples and carried out the experiments for this study. M.P. performed data analysis. Curated traces were independently checked by C.N. to avoid human bias. M.P. and C.N. prepared functionalized glass slides for single-molecule FRET. M.P. and R.T. wrote the manuscript. R.T. conceived and supervised the work.
Funding
EC | European Research Council (ERC)
Robert Tampé
Deutsche Forschungsgemeinschaft (DFG) (CRC 1507/P18)
Robert Tampé
Deutsche Forschungsgemeinschaft (DFG) (GRK 1986/B4.7)
Robert Tampé
References
1.
Structure, function, and evolution of bacterial ATP-binding cassette systems
Microbiol Mol Biol Rev
72
:317–364
PubMed
Google Scholar
2.
ABC transporters: the power to change
Nat Rev Mol Cell Biol
10
:218–227
PubMed
Google Scholar
3.
Structural and mechanistic principles of ABC transporters
Annu Rev Biochem
89
:605–636
PubMed
Google Scholar
4.
Mechanistic diversity in ATP-binding cassette (ABC) transporters
Nat Struct Mol Biol
23
:487–493
PubMed
Google Scholar
5.
Structural and functional diversity calls for a new classification of ABC transporters
FEBS Lett
594
:3767–3775
PubMed
Google Scholar
6.
Revisiting the role of ABC transporters in multidrug-resistant cancer
Nat Rev Cancer
18
:452–464
PubMed
Google Scholar
7.
The ABCs of immunology: structure and function of TAP, the transporter associated with antigen processing
Physiology (Bethesda)
19
:216–224
PubMed
Google Scholar
8.
Subnanometre-resolution electron cryomicroscopy structure of a heterodimeric ABC exporter
Nature
517
:396–400
PubMed
Google Scholar
9.
Asymmetric ATP hydrolysis cycle of the heterodimeric multidrug ABC transport complex TmrAB from Thermus thermophilus
J Biol Chem
286
:7104–7115
PubMed
Google Scholar
10.
Crystal structure and mechanistic basis of a functional homolog of the antigen transporter TAP
Proc Natl Acad Sci U S A
114
:E438–E447
PubMed
Google Scholar
11.
Conformation space of a heterodimeric ABC exporter under turnover conditions
Nature
571
:580–583
PubMed
Google Scholar
12.
A single power stroke by ATP binding drives substrate translocation in a heterodimeric ABC transporter
eLife
9
:e55943
PubMed
Google Scholar
13.
De novo macrocyclic peptides dissect energy coupling of a heterodimeric ABC transporter by multimode allosteric inhibition
eLife
10
:e67732
PubMed
Google Scholar
14.
Thermodynamic basis for conformational coupling in an ATP-binding cassette exporter
J Phys Chem Lett
11
:7946–7953
PubMed
Google Scholar
15.
Conformational coupling and trans-inhibition in the human antigen transporter ortholog TmrAB resolved with dipolar EPR spectroscopy
J Am Chem Soc
140
:4527–4533
PubMed
Google Scholar
16.
Reliability and accuracy of single-molecule FRET studies for characterization of structural dynamics and distances in proteins
Nat Methods
20
:523–535
PubMed
Google Scholar
17.
Precision and accuracy of single-molecule FRET measurements-a multi-laboratory benchmark study
Nat Methods
15
:669–676
PubMed
Google Scholar
18.
Single-molecule fluorescence resonance energy transfer in molecular biology
Nanoscale
8
:19928–19944
PubMed
Google Scholar
19.
Single-molecule FRET of membrane transport proteins
ChemBioChem
22
:2657–2671
PubMed
Google Scholar
20.
FRET-based dynamic structural biology: Challenges, perspectives and an appeal for open-science practices
eLife
10
:e60416
PubMed
Google Scholar
21.
Single-molecule FRET for probing nanoscale biomolecular dynamics
Nat Rev Phys
6
:587–605
Google Scholar
22.
Characterization of the kinetic cycle of an ABC transporter by single-molecule and cryo-EM analyses
eLife
9
:e56451
PubMed
Google Scholar
23.
CFTR function, pathology and pharmacology at single-molecule resolution
Nature
616
:606–614
PubMed
Google Scholar
24.
Conformational dynamics of the ABC transporter McjD seen by single-molecule FRET
EMBO J
37
:e100056
PubMed
Google Scholar
25.
Single-molecule dynamics reveal ATP binding alone powers substrate translocation by an ABC transporter
BioRxiv
:2025.2011.2027.690960
Google Scholar
26.
Mechanistic determinants of the directionality and energetics of active export by a heterodimeric ABC transporter
Nat Commun
5
:5419
PubMed
Google Scholar
27.
Substrate recognition diversity and transport dynamics of ABCC1
Nat Commun
16
:10499
PubMed
Google Scholar
28.
Cyanine fluorophore derivatives with enhanced photostability
Nat Methods
9
:68–71
PubMed
Google Scholar
29.
Leveraging Baird aromaticity for advancement of bioimaging applications
J Phys Org Chem
36
PubMed
Google Scholar
30.
A toolkit and benchmark study for FRET-restrained high-precision structural modeling
Nat Methods
9
:1218–1225
PubMed
Google Scholar
31.
Photophysics of fluorescent probes for single-molecule biophysics and super-resolution imaging
Annu Rev Phys Chem
63
:595–617
PubMed
Google Scholar
32.
Accurate distance determination of nucleic acids via Förster resonance energy transfer: implications of dye linker length and rigidity
J Am Chem Soc
133
:2463–2480
PubMed
Google Scholar
33.
The orientational freedom of molecular probes. The orientation factor in intramolecular energy transfer
Biophys J
26
:161–193
https://doi.org/10.1016/s0006-3495(79)85243-1
PubMed
Google Scholar
34.
Kappa-squared: from nuisance to new sense
J Biotechnol
82
:181–196
https://doi.org/10.1016/s1389-0352(01)00037-x
PubMed
Google Scholar
35.
DeepFRET, a software for rapid and automated single-molecule FRET data classification using deep learning
eLife
9
:e60404
PubMed
Google Scholar
36.
Alternating-laser excitation: single-molecule FRET and beyond
Chem Soc Rev
43
:1156–1171
PubMed
Google Scholar
37.
Single-molecule FRET imaging of GPCR dimers in living cells
Nat Methods
18
:397–405
PubMed
Google Scholar
38.
Reliable state identification and state transition detection in fluorescence intensity-based single-molecule Förster Resonance Energy-Transfer data
J Phys Chem B
122
:6134–6147
PubMed
Google Scholar
39.
Quantitative FRET studies and integrative modeling unravel the structure and dynamics of biomolecular systems
Curr Opin Struct Biol
40
:163–185
PubMed
Google Scholar
40.
Membrane interactions of Cy3 and Cy5 fluorophores and their effects on membrane-protein dynamics
Biophys J
119
:24–34
PubMed
Google Scholar
41.
Single-molecule FRET at 10 MHz count rates
J Am Chem Soc
146
:3539–3544
PubMed
Google Scholar
42.
Triple labeling resolves a GPCR intermediate state by using three-color single molecule FRET
J Am Chem Soc
147
:17689–17700
PubMed
Google Scholar
43.
Four-color alternating-laser excitation single-molecule fluorescence spectroscopy for next-generation biodetection assays
Clin Chem
58
:707–716
PubMed
Google Scholar
44.
Single cell-like systems reveal active unidirectional and light-controlled transport by nanomachineries
ACS Nano
15
:6747–6755
PubMed
Google Scholar
45.
Surface passivation for single-molecule protein studies
J Vis Exp
50549
PubMed
Google Scholar
46.
Substrate-bound and substrate-free outward-facing structures of a multidrug ABC exporter
Sci Adv
8
:eabg9215
PubMed
Google Scholar
Article and author information
Author information
Cite all versions
You can cite all versions using the DOI
10.7554/eLife.110967
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2026,
Pečak et al.
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
views
0
downloads
0
citations
0
Views, downloads and citations are aggregated across all versions of this paper published by eLife.





