 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库ryanodine receptor gene triggers functional and developmental muscle properties and could be used to assess the impact of humanmutations
Abstract
The ryanodine receptor (RYR) genes encode evolutionarily conserved calcium release channels involved in a wide range of calcium-dependent biological processes. Here, we show that the sole
Drosophila
RYR gene (
dRyR
) functions in differentiated somatic and cardiac muscle as well as in developing embryonic myotubes. In the larval body wall muscles, dRyR protein localizes at the SR membranes, and
dRyR
knockdown adversely affects muscle contractility, suggesting its conserved role in calcium-triggered E-C coupling. After
dRyR
attenuation, sarcomere, and mitochondrial patterns are severely impaired, showing
dRyR
involvement in structural muscle properties. However,
dRyR
is also prominently expressed and functionally required in growing embryonic muscles.
dRyR
loss of function leads to myotube growth defects and thin myofiber phenotypes, while its overexpression induces myofiber splitting. Given the structural and functional conservation of
dRyR
, we used
Drosophila
to test the impact of one human
RYR1
variant of unknown significance (VUS). Larvae carrying
p.Met4881Ile RYR1
VUS showed impaired mobility and altered structural muscle properties reminiscent of those seen in
dRyR
knockdown, thus indicating it is likely pathogenic. Overall, we show that
Drosophila dRyR
plays a conserved role in setting muscle contractility and structural muscle features. Our findings underline the still under-investigated role of
dRyR
as a promyogenic factor and provide a first example of the impact assessment of a human
RYR1
VUS in
Drosophila
.
Introduction
The calcium ion (Ca
2+
) concentration gradient is known to be a crucial second messenger signal in all eukaryotic cells. The calcium release channels encoded by ryanodine receptor genes are essential to maintaining correct Ca
2+
dynamics across biological membranes. RYR proteins are cellular sites of interactions with ryanodine, an alkaloid isolated from the stem wood of the plant
Ryania speciosa
(
Jenden and Fairhurst, 1969
) that was used for several decades as an insecticide. Because of its muscle-paralyzing effect in humans, ryanodine has been replaced by insect-specific derivatives that are non-toxic for mammals. Analysis of the RYR multi-domain structures and phylogenetic relations among different taxa yields a model suggesting that it evolved from inositol 1,4,5-trisphosphate receptor (IP
3
R)-like ancestral Ca
2+
release channels. The RYR activity is strictly controlled and may respond to the presence of ions (Ca
2+
, Mg
2+
, and Zn
2+
), proteins (calmodulin (Cam), and FK-506 binding protein (FKBP12/12.6)), and small molecules, such as ATP, caffeine, and ryanodine. The domain structure is highly conserved. For example, the SPIa kinase and ryanodine receptor (SPRY) domain shares high sequence identity between vertebrates and invertebrates and is engaged in protein-protein interactions with several protein families (
Hadiatullah et al., 2022
).
The number of RYR copies ranges across taxa. Mammalian genomes carry three RYR genes. For example, in humans,
RYR1
,
RYR2,
and
RYR3
are located on chromosomes 19q13.2, 1q43, and 15q13.3–14, respectively. Non-mammalian vertebrates, such as
Xenopus laevis
and chicken, have two
RYR
copies (
Ottini et al., 1996
) while in a zebrafish genome, there are five genes:
RYR1a, RYR1b, RYR2a, RYR2b
, and
RYR3
displaying high similarity to other vertebrate RYR genes (
Wu et al., 2011
). The greater number of RYR genes in zebrafish than in mammals and birds is thought to result from teleost-specific whole-genome duplication (
Howe et al., 2013
;
Postlethwait et al., 1998
). In the invertebrates
Drosophila melanogaster
,
Caenorhabditis elegans,
live scallop (
Placopecten magellanicus
), and lobster (
Homarus americanus
), a single RYR gene has been identified (
Hasan and Rosbash, 1992
;
Maryon et al., 1996
;
Murayama and Kurebayashi, 2011
;
Quinn et al., 1998
;
Xu et al., 2000
). However, through alternative splicing, the single invertebrate RYR gene produces several isoforms, thus increasing the diversity of the available protein pool (
De Mandal et al., 2019
;
George et al., 2007
).
It is well known that RYR genes are expressed in muscle cells and play a crucial role in muscle contraction, which results from excitation-contraction (E-C) coupling, a series of events involving the conversion of electrical stimulus to Ca-dependent mechanical response. However, RYRs are also expressed in many other animal tissues, including the central nervous system (
Klatt Shaw et al., 2018
;
Liu et al., 2005
) and are involved in housekeeping functions in the cells of the adult organism and in developmental processes (
Fill and Copello, 2002
).
RYR1
mutations underlie an array of diseases, including muscle-impairing central core disease (CCD), one of the most frequent congenital myopathies, and malignant hyperthermia susceptibility (MHS), characterized by severe reaction to anesthetics, excessive heat, or exercise, which can be fatal. In addition, several
RYR2
mutations have also been implicated in cardiopathic catecholaminergic polymorphic ventricular tachycardia (CPVT) and arrhythmogenic right ventricular dysplasia of type 2 (ARVD2) (
Lanner, 2012
). Of the numerous mutations identified to date in RYR genes, most are missense mutations with single amino acid substitutions. However, several deletions, duplications, and frame shift mutations have also been identified (
Lanner, 2012
).
Here, we analyze the phylogenetic origin, expression and function of the
Drosophila
dRyR gene. Our data show that
dRyR
is not only required for contractile properties of differentiated somatic and cardiac
Drosophila
muscle but also influences muscle structure and plays an instructive role in muscle development. Structural muscle changes in a
dRyR
loss-of-function context suggest its implication in age-associated muscle decline, while severe developmental muscle defects observed in
dRyR
mutant embryos provide insights into early-onset RYR-related myopathies. Considering extensive structural and functional
dRyR
conservation, we generated a
Drosophila
model of one undiagnosed human
RYR1
variant mutation (
p.M4881Ile
) and found that it negatively impacted muscle structure and function, making it likely pathogenic. We report a detailed functional analysis of
dRyR
. Our findings pinpoint the under-investigated role of
dRyR
in embryonic muscle development and demonstrate that
Drosophila
could be used for assessing impacts of human
RYR1
variant mutations of unknown significance.
Results
Drosophila dRyR
belongs to the RYR gene family and shows conserved somatic and cardiac muscle-associated expression and function
dRyR
is the sole RYR gene family member in
Drosophila
. We applied the maximum likelihood method and the Kimura 2-parameter model (
Kimura, 1980
) to infer the evolutionary history of
dRyR
. Based on evolutionary distance studies,
RYR2
is the most ancient of the three human RYR genes (
Ding et al., 2017
) and shows the highest sequence homology with
Drosophila dRyR
. We compared sequences of
RYR2
from selected mammalian and non-mammalian vertebrates with the sequence of single invertebrate RYR, including fruit fly
dRyR
to illustrate its phylogenetic origin (
Figure 1A
;
Ding et al., 2017
;
Mackrill, 2012
;
McKay and Griswold, 2014
;
Takeshima et al., 1994
). Evolutionary analyses were conducted in MEGA X (
Kumar et al., 2018
;
Stecher et al., 2020
). We observed that single invertebrate RYR genes, here from
Drosophila
and
C. elegans
, clustered into a separate branch connected to the vertebrate
RYR2
branch that clustered 100% (
Figure 1A
). The vertebrate
RYR2
branch was then subdivided clonally from
Xenopus
through zebrafish, chick, and mouse to human. All clonal branches in vertebrates clustered 100% (
Figure 1A
), indicating that
RYR2
genes are closely related and most probably evolved from a single invertebrate RYR.
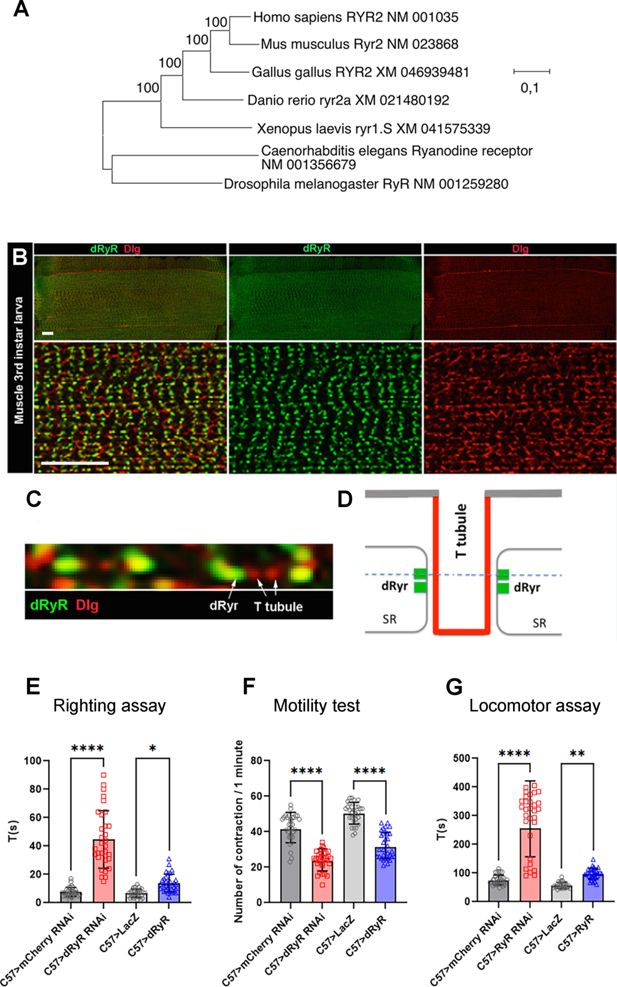
Phylogenetic origin of
Drosophila
RYR (dRyR), its body wall muscle associated expression and role in locomotion.
(
A
) Evolutionary analysis by the maximum likelihood method of ryanodine receptor genes (RYR) from different taxa. The tree with the highest log likelihood (−95718.65) is shown. The percentage of trees in which the associated taxa clustered is shown next to the branches. (
B
) A wide view (upper panels) and a zoomed view (lower panels) of ventral VL3 larval muscle stained for dRyR (green) and discs large (Dlg) (red) that labels T-tubules. (
C
) A high-magnification view showing dRyR dots at the interface of T-tubules (arrows). (
D
) Scheme presenting subcellular location of dRyR receptor at the sarcoplasmic reticulum (SR) membrane in a close vicinity of T-tubules. The dotted line refers to the optical level of confocal view in (
C
). (
E–G
) Larval muscle targeted
dRyR
knockdown (C57>dRyRRNAi) leads to a marked decline in muscle performance compared to control (C57>mCherryRNAi). Three muscle performance tests were applied: (
E
) righting test, (
F
) motility test, and (
G
) locomotor test. Overexpression of
dRyR
in larval muscle (C57>dRyR) impacts muscle performance measured by the locomotor and motility tests (
F, G
). Scale bar: 20 μm. All statistical analyses were performed using Prism. The one-way ANOVA test was used for comparisons of datasets. Bar plot represent the mean and the standard deviation. On the figures, statistical comparisons of sample vs control are indicated as ****
p
≤0.0001; ***
p
≤0.001; **
p
≤0.01; *
p
≤0.05; ns>0.05.
Figure 1—source data 1
The source data for three larval muscle performance tests: righting aasay, motility test and locomotor assay.
Download elife-111053-fig1-data1-v1.xlsx
Expression of RYR genes in vertebrates has been extensively studied and described in various excitable cells, including skeletal and cardiac muscles and neurons, and in non-excitable cells, such as pancreatic beta cells and lymphocytes (Rossi & Sorrentino, 2002). By contrast, in spite of early works by
Hasan and Rosbash, 1992
and
Sullivan et al., 2000
no systematic analyses have yet been performed to assess the developmental expression pattern of the sole
Drosophila dRyR
gene.
We first tested the expression of
dRYR
at protein level. In differentiated body wall muscle of third instar larva (
Figure 1B and C
)
dRyR
was detected in a discrete striated pattern (
Figure 1B
), which in a zoom view revealed highly ordered punctate dRyR protein localization in close vicinity to discs large (Dlg)-positive T-tubules (
Figure 1C
and scheme in
Figure 1D
). Thus, in
Drosophila
muscle, like in vertebrates, dRyR localizes at the T-tubule interface, a sub-cellular localization consistent with its calcium release role at SR and E-C coupling function during muscle contraction.
We also assessed muscle-associated expression of
dRyR
transcripts (Fig. S1). Of ten
dRyR
transcript isoforms, all coded for proteins of similar amino acid (aa) length, ranging from 5113–5134 aa and molecular weight about 580 kDa (
). To test
dRyR
isoform expression, we applied the FISH-HCR technique (
Choi et al., 2016
) and four probes targeting alternative exons 10, 11, 22, and 23 (
Figure 1—figure supplement 1A
). In this setup we detected the expression of the A, B, F, G, H, J isoforms with the
dRyR
Ex10 probe, A, B, C, D, E, I isoforms with the Ex11 probe, A, B, C, D, E, F, G isoforms with the Ex22 probe and H, I, J isoforms with the Ex23 probe (
Figure 1—figure supplement 1A
). FISH-HCR experiments performed on differentiated third instar larval muscles revealed muscle-associated specific signals with all four probes (
Figure 1—figure supplement 1B-E
).
dRyR
transcripts were detected at the periphery of nuclei, in the sarcoplasm and in a repeated striated pattern following sarcomeres. Similar signals observed with the four probes indicated that at least the isoforms A and B that are commonly targeted by the Ex10, Ex11, and Ex22 probes and one of the isoforms H, I, or J targeted by the Ex23 probe were present in the functional body wall muscle. We could not, however, rule out the possibility that most or even all
dRyR
isoforms were expressed in the differentiated body wall muscles. Supporting this, we note that nine out of ten
dRyR
isoforms (isoform I being an exception) share the same transcription start site, suggesting that a common core promoter regulates their expression.
Detected muscle-associated expression of
dRyR
prompted us to test its involvement in muscle contraction and larva mobility. We observed that muscle-targeted attenuation of
dRyR
expression resulted in reduced muscle performance with a significantly longer time required for larvae to switch from the dorsal to the ventral position (
Figure 1E
– righting test) and a reduced number of muscle contraction waves compared to control recorded in 1 min (
Figure 1F
– motility test).
Finally,
dRyR-RNAi
larvae crawled inefficiently and were unable to move over a longer distance (
Figure 1G
– locomotor test). In parallel, overexpressing
dRyR
in larval muscles also impaired muscle function, with fewer contraction waves in 1 min compared to control and a slower locomotion (
Figure 1F and G
), while the time required to switch from the dorsal to the ventral position remained unchanged (
Figure 1E
).
Altogether, our data extend previous observations of affected muscle contractility in RyR mutants (
Sullivan et al., 2000
) and suggest that
Drosophila dRyR
, like its vertebrate counterparts, ensures correct muscle function, likely acting as a sarcoplasmic reticulum (SR) calcium release channel essential for muscle contraction and E-C coupling.
Given that vertebrate RYR genes (
RYR2
in humans) also play an instrumental role in cardiac muscle function, we sought to determine whether dRyR protein could be detected in the fly heart and whether it could influence heartbeat variables. In the adult
Drosophila
heart, dRyR protein was detected predominantly in the circular muscle fibers ensuring cardiac contractions (
Figure 2A–A
), which appear structurally affected (
Figure 2C and E
) in a heart-specific
dRyR RNAi
context (Hand >dRyR RNAi). Consistent with this, heart-targeted attenuation of
dRyR
resulted in an abnormal M-mode heart profile (
Figure 2B and D
) associated with a longer heart period (
Figure 2F
) and a slow heart rate (
Figure 2G
), with significantly increased diastolic interval (
Figure 2H
). Attenuation of cardiac
dRyR
expression also led to arrhythmic heartbeat (
Figure 2J
), increased systolic diameter (
Figure 2K
) and reduced fractional shortening (
Figure 2M
). As the diastolic diameter remained unchanged, we conclude that cardiac dRyR knockdown affects cardiac performance without causing dilated cardiomyopathy. In contrast to dRyR-RNAi, increasing
dRyR
cardiac expression had only a minor influence on cardiac variables (
Figure 2F–M
) with increased systolic diameter (
Figure 2K
) but no effect on cardiac contractility (
Figure 2M
). Overall, observed
dRyR
loss-of-function adult fly heart phenotypes with a slow heart rate and increased arrhythmia correlate with impaired cardiac function in RyR mutant larvae (
Sullivan et al., 2000
). We hypothesize that dRyR RNAi-induced impairment of Ca
2 +
homeostasis could contribute to cardiac aging, for which
Drosophila
is a recognized model (
Nishimura et al., 2011
).
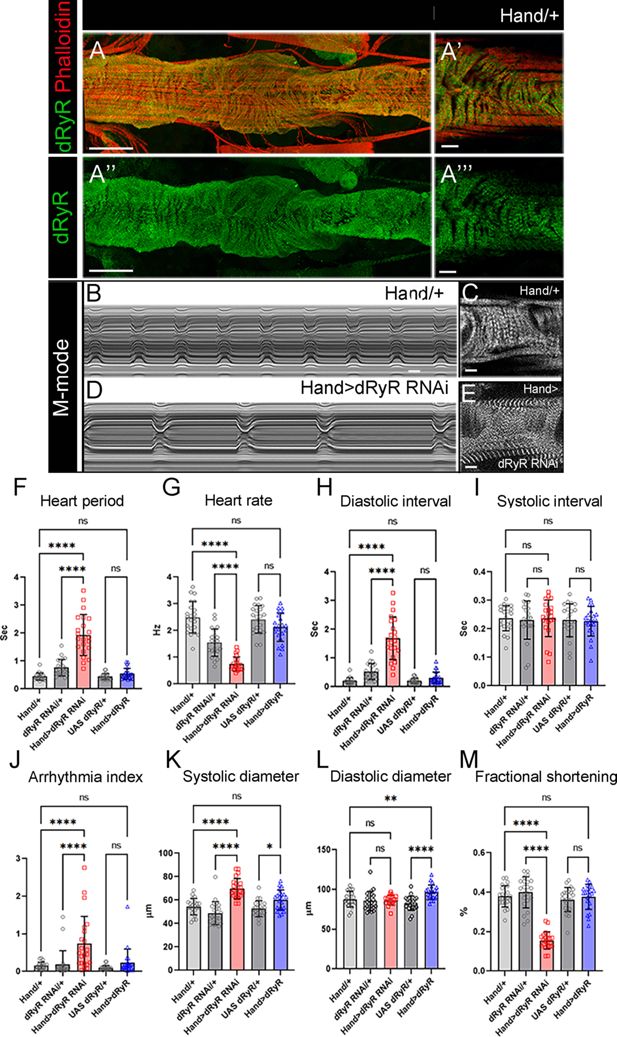
Drosophila
RYR (dRyR) is expressed in the heart tube and is required for correct heartbeat.
(
A-A”’
) Adult heart tube labeled for dRyR (green) and actin (red). (
A’, A’’’
) Zoomed views revealing
dRyR
expression in circular fibers. (
B,D
) M-modes of control Hand/+ (
B
) and Hand >dRyR RNAi (
D
) contexts showing a slow heart rate induced by
dRyR
attenuation. Compared with control (
C
), circular fibers in Hand >dRyR RNAi (
E
) context showed a fuzzy pattern suggesting an affected sarcomeric organisation. (
F–M
) Heartbeat variables in cardiac
dRyR
knockdown (Hand >dRyR RNAi) and cardiac
dRyR
overexpression contexts (Hand >dRyR). Scale bar: 50 μm in A, A’’; 10 μm in A’, A’’’, C, E. All statistical analyses were performed using Prism. The one-way ANOVA test was used for comparisons of datasets. Bar plot represent the mean and the standard deviation. On the figures, statistical comparisons of sample vs control are indicated as ****
p
≤0.0001 *
p
≤0.05; ns>0.05.
Figure 2—source data 1
Source data for heartbeat variables in cardiac dRyR knockdown (Hand >dRyR RNAi) and cardiac dRyR overexpression contexts (Hand >dRyR).
Download elife-111053-fig2-data1-v1.xlsx
In view of the reduced motility of third instar larva with the attenuated
dRyR
(
Figure 1E–G
), we examined whether structural properties of body wall muscles were adversely affected. We first found that muscle-targeted attenuation of
dRyR
(C57 >dRyR RNAi) led to a significantly reduced larva body length (
Figure 3B and Q
) compared to control (
Figure 3A and Q
), an observation that correlates with previously observed (
Sullivan et al., 2000
) reduced body size of
dRyR
16
mutant larvae. Though to a lesser extent, the overexpression of
dRyR
in body wall muscles also impacted larva length (
Figure 3D and Q
). These changes in larva size in loss and gain of
dRyR
function correlated with a reduced longitudinal muscle length (
Figure 3E–H and R
), which in turn correlated with shortening of Kettin/D-Titin-labelled sarcomeres (
Figure 3I–L and T
) and reduced number of myonuclei (
Figure 3E–H and S
). Because RYR-mediated calcium homeostasis involves dynamic interactions between the sites of calcium release from the SR and calcium uptake by the mitochondria (
Li et al., 2025
), we examined whether in
Drosophila dRyR
loss and/or
dRyR
gain of function could adversely affect mitochondria pattern in the larval muscles. We noted that the I band-associated striated mitochondria pattern was lost in the C57 >dRyR RNAi context (
Figure 2—figure supplement 1B
) and appeared irregular when
dRyR
was overexpressed in muscles (
Figure 2—figure supplement 1D
) compared to the wild-type larval muscles (
Figure 2—figure supplement 1A, B and E
).
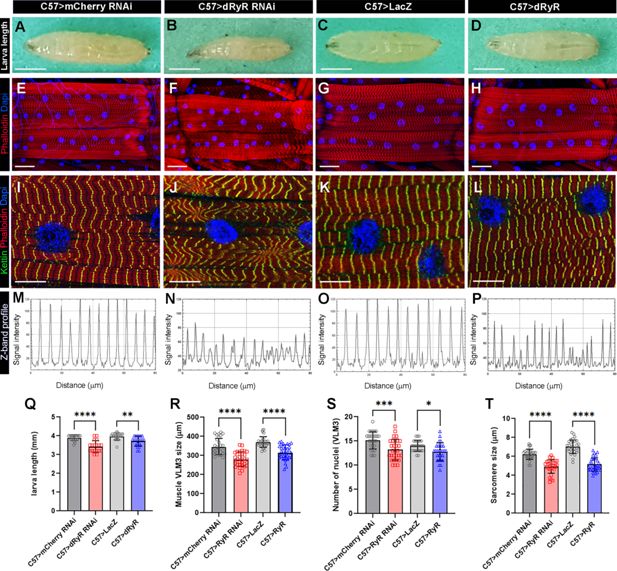
Muscle targeted
Drosophila
RYR (dRyR) loss and gain of function impacts body size and structural muscle properties.
(
A–D
) General views of third instar larva in control (
A, C
), muscle-targeted
dRyR
attenuation (
B
) or overexpression (
D
). (
E–H
) Representative VL3 and VL4 ventral longitudinal muscle views from age-matched third instar larvae in control (
E, G
), C57>dRyR RNAi (
F
), and C57>dRyR (
H
) contexts. Muscle fibers and nuclei were revealed with phalloidin (red) and DAPI (blue), respectively. (
I–L
) Zoomed views of VL3 muscles of control (
I, K
), C57>dRyR RNAi (
J
), and C57>dRyR (
L
) larvae triple-stained for phalloidin (red), DAPI (blue), and Kettin/D-Titin (green). (
M–P
) Z band profiles (Kettin signal intensity plot) from zoomed views of VL3 muscles presented in (
I–L
). (
Q
) Statistical representation of third instar larva length. (
R–T
) Statistical representation of VL3 muscle characteristics: (
R
) VL3 muscle length; (
S
) number of nuclei; and (
T
) sarcomere size. Scale bar: 1 mm in A-C; 50 μm in D-F; 20 μm in G-I. Bar plots represent the mean and the standard deviation. All statistical analyses were performed using Prism. The one-way ANOVA test was used for comparisons of datasets. Bar plot represent the mean and the standard deviation. On the figures, statistical comparisons of sample vs control are indicated as ****
p
≤0.0001; ***
p
≤0.001; **
p
≤0.01; *
p
≤0.05; ns>0.05.
Figure 3—source data 1
Source data for third instar larva length (Q) and VL3 muscle characteristics: (R) VL3 muscle length; (S) number of nuclei; and (T) sarcomere size.
Download elife-111053-fig3-data1-v1.xlsx
Both muscle-targeted attenuation and gain of
dRyR
function led not only to impaired muscle functions but also to overall reduction of muscle size and myofibrillar disarray associated with a downsizing of sarcomeres and mitochondrial mismatch. Altered structural muscle features observed in
dRyR
-attenuated
Drosophila
larvae are reminiscent of myofibrillar and mitochondrial pattern defects reported in mice harboring a pathogenic
RYR1
mutation (
Elbaz et al., 2019
).
dRyR
is expressed during embryonic muscle development and is required for correct myogenic differentiation
Previous reports provide evidence that RYR-dependent elevation of intracellular calcium promotes late steps of myogenic differentiation and, in particular, fusion of myoblasts to myotubes (
Eigler et al., 2021
;
Sinha et al., 2022
). In parallel, a recent study on RYR1-depleted primary myoblasts revealed the calcium-independent inhibitory role of RYR1 in myogenic differentiation (
Tourel et al., 2025
). To further explore myogenic roles of RYRs, we tested
dRyR
expression and function during embryonic development.
Sullivan et al., 2000
reported embryonic
dRyR
transcript expression in body wall and visceral muscle precursors. Here, we tested dRyR protein expression and found that it was prominently expressed in the mesodermal derivatives in embryos. We detected dRyR protein in the developing visceral, somatic, and cardiac muscle cells (
Figure 4
). Regarding body wall muscles, dRyR could be detected in the somatic muscle precursors starting from embryonic stage 12 (
Figure 4A and A
), accumulated in the growing myotubes at mid-stage embryos (
Figure 4B and B
) and continued to be expressed in the developing muscle fibers at later embryonic stages (
Figure 4C–D
). At embryonic stage 16, dRyR protein was distributed in a discrete granular pattern within the cytoplasm of myofibers and appeared excluded from the myonuclei (
Figure 4D and D
). Thus, dRyR protein was detected from the early phase of myogenic differentiation that encompasses specification of muscle founders and first myoblast fusion events and continues during the second phase of fusion and myofiber growth and maturation. We also applied HCR-FISH to test
dRyR
transcript isoform expression in the developing somatic muscle (
Figure 3—figure supplement 1
). We found that
dRyR
A, B, F, and G isoforms harboring alternative exons 10 and 22 were actively transcribed in the developing muscle, whereas the remaining
dRyR
isoforms were barely detected (
Figure 3—figure supplement 1
).
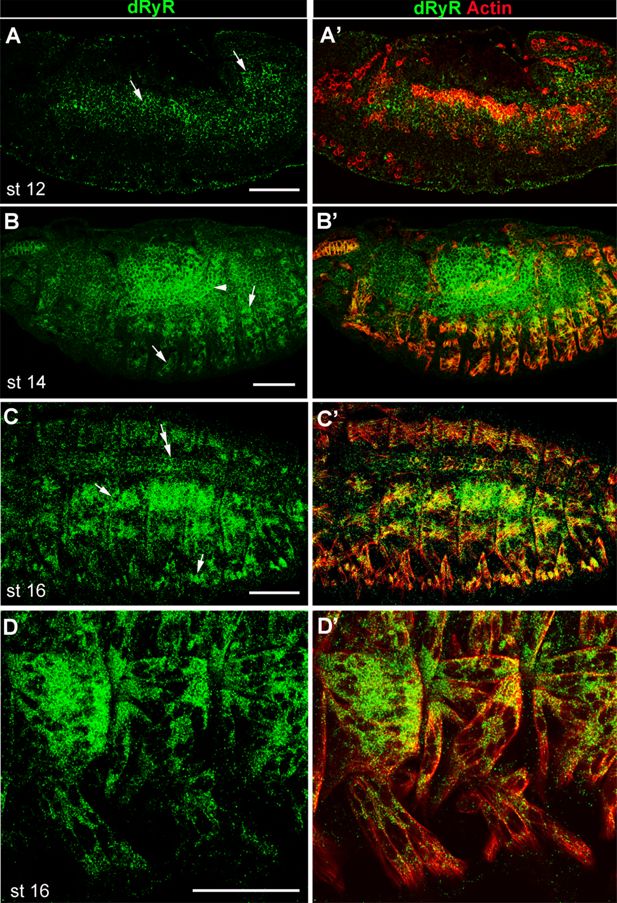
Developmental
Drosophila
RYR (dRyR) protein pattern in embryos.
(
A-A’
) lateral view of a stage 12 embryo. dRyR (green) could be detected in the somatic and visceral muscle precursors (arrows in A) also revealed by Actin (red) (
A’
). (
B-C’
) dorso-lateral views of stage 14 (
B,B’
) and stage 16 (
C,C’
) embryos. dRyR accumulates in body wall muscle precursors (arrows in B and C) and in visceral muscle of the midgut (arrowhead in B) and in the dorsally aligned cardioblasts (double-head arrow in C). (
D,D’
) Subcellular dRyR pattern in ventral muscle precursors at embryonic stage 16. Note granular cytoplasmic distribution of dRyR. Scale bar: 50 μm.
Previous analyses (
Sullivan et al., 2000
) showed that muscle contraction was compromised in larvae carrying a hypomorphic
dRyR
16
mutant allele.
dRyR
mutant larvae were also smaller in size and died before the pupation stage. However, whether
dRyR
embryonic expression has a functional impact on larval muscle development has not yet been assessed. Accordingly, we analyzed embryonic muscle pattern in late-stage
dRyR
16
mutant embryos (
Figure 5
). We observed a wide range of developmental somatic muscle defects with predominant phenotype of thin myofibers present in 64% of abdominal A2-A5 hemisegments and more severely sphere-shaped (arrowheads in
Figure 5B
) or missing myofibers (asterisks in
Figure 5B
) observed in 25% of hemisegments. In addition, in 10% of hemisegments with
dRyR
loss of function led to supernumerary lateral transverse (LT) muscles (arrows in
Figure 5B
), a phenotype that could arise from LT muscle splitting (
Bertin et al., 2021
). Predominant thin/misshaped/missing myofiber phenotype in a
dRyR
loss-of-function context suggests a pro-myogenic role during development. To further characterize the role of dRyR during myogenesis, we analyzed embryonic LT muscle phenotypes in LT-targeted
dRyR
attenuation and gain-of-function contexts (
Figure 5C–E
). Like in
dRyR
16
mutant embryos,
dRyR
RNAi knockdown in LT muscles resulted in thin and/or misshaped LT muscles (
Figure 5D and H
) observed in 88% of hemisegments and in rare cases of LT splitting, found in 8% of hemisegments (
Figure 5D and H
). We noted no loss of LT muscles in
dRyR
RNAi embryos. Consistent with the promyogenic role of
dRyR
, LT-targeted overexpression of
dRyR
appears to promote LT splitting phenotype (
Figure 5E and H
) found in 18% of hemisegments. Calmodulin Cam is the major calcium-dependent RYR regulator. The Ca
2+-
bound Cam at high calcium levels acts as RYR inhibitor (
Fruen et al., 2003
). We thus tested effects of Cam attenuation in LTs and found that it results in a dRyR overexpression-like phenotype (
Figure 5F and H
). This suggests that during myogenesis Cam is present mainly in a calcium-bound form that represses dRyR. Another major regulator of calcium homeostasis, the endoplasmic reticulum calcium pump SERCA is required to maintain high calcium levels in the ER lumen (
Suisse and Treisman, 2019
). To test its role in myogenesis we analysed Lms >SERCA RNAi embryos. We observe that SERCA-depleted LT muscles display growth defects with predominant thin myofiber phenotype (
Figure 5G and H
) also observed in Lms >dRyR RNAi context (
Figure 5D and H
). Thus, both dRyR-regulated cytosolic and SERCA-regulated ER lumen calcium levels are required to promote muscle development.
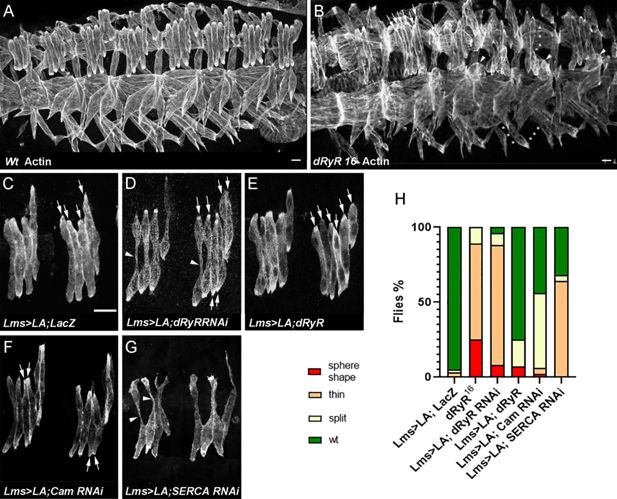
Drosophila
RYR (dRyR) is required for correct embryonic muscle development.
(
A, B
) ventro-lateral views of stage 16 embryos stained for actin to reveal embryonic muscle pattern in wild-type (
A
) and in homozygous
dRyR
16
mutant embryo (
B
). Note a wide range of developmental muscle defects that could be observed in
dRyR
loss-of-function context. Asterisks in B pinpoint muscle fiber loss, arrowheads indicate the myofibers that failed to extend and remained as myospheres and a series of arrows point to supernumerary lateral transverse myofibers (6 instead of 4). (
C–E
) Effects of lateral transverse (LT) muscle-targeted attenuation (
D
) and overexpression (
E
) of
dRyR
. Lateral transverse (LT) muscles were revealed by targeted expression of LifeActinGFP (LA) transgene using LT-specific Lms-GAL4 driver. (
C
) Four LT muscles (arrows) are seen in a control Lms >LA;LacZ context. (
D
) dRyRRNAi attenuation led to misshaped thin LTs (arrowheads) – major phenotype and to an occasional LT muscle split phenotype (6 LTs indicated by arrows). (
E
) LT targeted overexpression of
dRyR
resulted mainly in LT muscle splitting (arrows). (
F
) Cam attenuation induced mostly LT muscle splitting (arrows) while (
G
) SERCA RNAi knockdown lead to affected myofiber growth with thin LT muscle phenotype (arrowheads). (
H
) Statistical representation of LT muscle phenotypes in
dRyR
mutants and LT targeted
dRyR
knockdown, gain-of-function, and Cam and SERCA attenuation contexts. The statistical analyses were performed using Prism - contingency test; 50–60 segments/genotype. Scale bar: 10 μm.
Figure 5—source data 1
Source dat for LT muscle phenotypes in dRyR mutants and LT targeted dRyR knockdown, gain-of-function, and Cam and SERCA attenuation contexts.
Download elife-111053-fig5-data1-v1.xlsx
Interestingly, live imaging (
Figure 4—figure supplement 1
) shows that the sphere-shaped muscle phenotype arises from the impaired LT myotube extension and not from retraction of already extended myotubes. Also, myonuclei remained on LT extremities and did not spread along the abnormally thin myofibers. The reduced number of myonuclei (2–4 per dRyR RNAi LT myofiber (
Figure 4—figure supplement 1B
, lower panel) instead of 4–6 in control LTs (
Figure 4—figure supplement 1A
lower panel)) points to the role of
dRyR
in the second wave of fusion (
Eigler et al., 2021
). This observation is consistent with the fact that overexpression of
dRyR
induced the LT split phenotype (
Figure 4E
) known to be promoted by an excessive myoblast fusion (
Bertin et al., 2021
). Because the embryonic LT muscle defects in
dRyR
-mutant embryos are associated with a reduced calcium signal in LTs (
Figure 4—figure supplement 1C
, D), we hypothesize that
dRyR
acts as a promyogenic factor ensuring correct calcium levels in the developing myotubes.
Assessing the impact of the
RYR1
undiagnosed variant mutation in
Drosophila
Drosophila
dRyR shares 45% aa sequence identity with human RYR1 and RYR2, and all protein domains are conserved, with up to 75% of identity for the most C-terminal RIH domain (
Figure 5—figure supplement 1A
). Thus, although the 3D conformation of the
Drosophila
dRyR has not yet been established, high sequence and positional conservation of functional domains suggest that the conformation of dRyR protein is similar to that of its vertebrate counterparts. Also, the distribution of pathogenic mutations identified in human
RYR1
and
RYR2
genes (reviewed by
Lanner et al., 2010
), clustered in three hot spots, correlates with the positions of conserved domains. As revealed by the identity heat map (
Figure 5—figure supplement 1B
), the hot spot regions of human
RYR1 and RYR2
mutations align with the most conserved portions of the
Drosophila
dRyR indicating suitability of the
Drosophila dRyR
for modeling human
RYR
gene mutations and their impact on muscular and cardiac systems.
Over the last decades, whole genome sequencing has identified large numbers of variant mutations within the
RYR
genes, most of which are classified as variants of unknown significance (VUS). The recessive
c.14643G>A
/p.Met4881Ile missense
RYR1
mutation was identified in a young patient with a phenotype of congenital myopathy with a delayed acquisition of motor function. At the histopathological level, filamentous aggregates were present in muscle biopsies. This very rare mutation for which a link with a muscle disorder has not yet been evaluated, is located in the
RYR1
region encoding calcium pore.
We made use of the conserved muscle function and structural similarity between dRyR and human RYRs (
Figure 5—figure supplement 1A
) demonstrated here to generate a
Drosophila
model of variant
p.Met4881Ile RYR1
and assess its impact on larval muscle function and structure (
Figure 6
). The
p.Met4881Ile RYR1
VUS-carrying larvae were homozygous-viable but were significantly smaller (
Figure 6B and I
) than control larvae (
Figure 6A and I
). They had shorter ventral longitudinal muscles (
Figure 6D and J
), which harbored fewer myonuclei (
Figure 6D and K
) and were characterized by shorter sarcomeres (
Figure 6F and H
) compared to control (
Figure 6C, E and G
). As revealed by the larva motility tests (
Figure 6M and O
) the
p.Met4881Ile RYR1
VUS impacted not only structural but also contractile muscle properties, leading to a reduced frequency of peristaltic body wall muscle contraction (
Figure 6N
), slower larva locomotion (
Figure 6O
), and compromised muscle performance revealed by the righting test (
Figure 6M
).
Figure 6
Download asset
Open asset
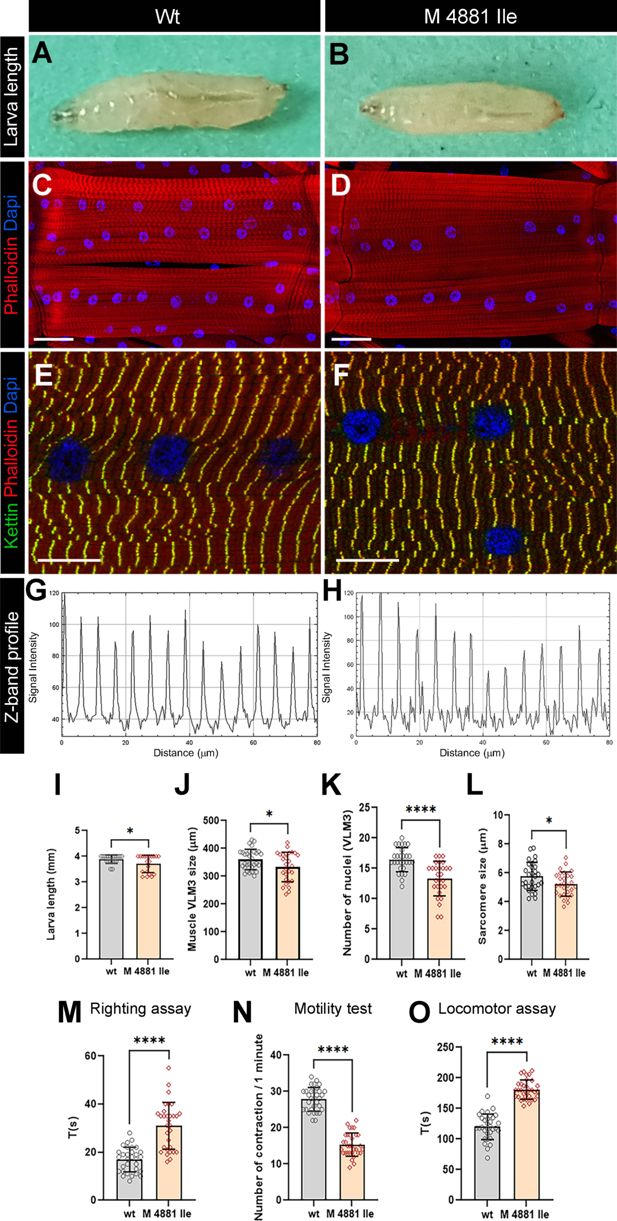
Modeling human RYR1 variant mutation in Drosophila
.
(
A,B
) Age-matched third instar wild-type (
A
) and
RyR1 p.Met4881Ile
mutant (
B
) larvae. Note a reduced size of larvae carrying
RYR1
variant mutation. (
C, D
) Representative views of ventral longitudinal (VL) muscles in wild-type (
C
) and
RYR1
variant mutant larvae (
D
). Note slightly reduced VL3 muscle length and reduced number of myonuclei in mutant condition. (
E–H
) Z band profile revealing reduction of sarcomere length in
RYR1
variant context (
F, H
) compared to control (
E, G
). Kettin is in green, Phalloidin in red, and DAPI in blue. (
I–L
) Statistical representation of larva length (
I
) and structural muscle variables (
J–L
) in wild-type and
p.Met4881Ile RYR1
variant mutation contexts. (
M–O
) Statistical assessment of functional larval muscle performance using righting test (
M
), motility test (
N
), and locomotor test (
O
) in wild-type and
RYR1
mutant conditions. Scale bar: 1 mm in A-B; 50 μm in C-D; 20 μm in E-F. All statistical analyses were performed using Prism. The
t
-test was used to compare control to variant context. Bar plot represent the mean and the standard deviation. On the figures, statistical comparisons of sample vs control are indicated as ****
p
≤0.0001; *
p
≤0.05.
Figure 6—source data 1
Source data for larva length (I), structural muscle variables (J–L) and for functional larval muscle performance using righting test (M), motility test (N), and locomotor test (O) in wild-type and RYR1 mutant conditions.
Download elife-111053-fig6-data1-v1.xlsx
These structural and functional muscle phenotypes are reminiscent of those observed in
dRyR RNAi
larvae, suggesting that
p.Met4881Ile
variant mutation negatively influences
dRyR
expression and/or function and could likely contribute to RYR-related myopathies in humans.
Discussion
The ryanodine receptors encoding calcium release channels are key regulators of intracellular calcium level, largely conserved among vertebrate and invertebrate species. By their involvement in many calcium-dependent biological processes, RYR genes are vital for normal cellular functions, while mutations in RYR genes underlie a broad spectrum of human diseases, including neuromuscular and cardiac disorders (
Marks, 2023
). Because of their large size (over 500 kDa), structural complexity, many isoforms, and broad spectrum of interacting factors, even though their role in promoting muscle contraction via E-C coupling is well-characterized, other RYR functions remain only partially understood. Here, we focus on
dRyR,
a single
Drosophila
member of the RYR gene family, and characterize its expression and function in differentiated and developing muscle.
Drosophila dRyR
shares 45% aa sequence identity with its human counterparts. Considering previous evolutionary distance studies (
Ding et al., 2017
), we show that the phylogenetic dRyR branch is connected to
RYR2
, the most ancient of the three human
RYRs
. Furthermore, a sequence alignment reveals an extensive conservation of all protein domains between
dRyR
and human RYRs, while a heatmap of conserved regions highlights previously identified hot spots of pathogenic
RYR1
and
RYR2
mutations. Remarkably, several aa residues at the functionally relevant channel pore domain (
Wang et al., 2012
) are conserved between
Drosophila
and humans.
Consistent with potential conservation of
dRyR
function in triggering Ca
2+
dynamics across ER membranes, we detected ER-associated punctate dRyR protein expression in differentiated body wall muscles. Our data extend previously reported analyses (
Sullivan et al., 2000
;
Vázquez-Martínez et al., 2003
) providing evidence that
dRyR
is not only expressed in visceral muscle and the nervous system but also prominently in differentiated striated somatic and cardiac muscle. We also wanted to know whether different
dRyR
transcript isoforms were differentially expressed in body wall muscles. However, we did not detect any such differential expression with four HCR-FISH probes targeting alternative
dRyR
exons. The fact that most
dRyR
transcript isoforms are present in differentiated larval muscles is consistent with their common transcription start sites. However, whether all
dRyR
isoforms are collectively required for larval muscle function requires further investigation. In line with muscle-associated
dRyR
expression and the excitation-contraction coupling role of its vertebrate orthologs, RNAi knockdown of
dRyR
leads to reduced muscle contractility and severely impaired larva mobility. Interestingly, in addition to impaired muscle function,
dRyR
attenuation causes extensive structural muscle defects, including reduced muscle size, smaller and aberrant sarcomeres, and degraded mitochondria pattern. All these functional and structural muscle defects are reminiscent of those of aged muscle, indicating that maintenance of
dRyR
-triggered calcium management could prevent muscle aging. This also applies to cardiac muscle, which in the
dRyR RNAi
context shows aging-associated slow heart rate and arrhythmia (
Blice-Baum et al., 2019
).
dRyR
transcripts and dRyR protein were also detected in the embryonic muscle precursors from embryonic stage 12 to late-stage embryos, indicating that
dRyR
could be involved in managing Ca
2+
levels throughout myogenesis. In contrast to differentiated larval muscles, HCR-FISH experiments show that only a subset of
dRyR
splice isoforms is expressed in the developing muscles. Our developmental
dRyR
expression data, focusing on somatic muscle, extend previous more general analyses of
dRyR
expression and function (
Takeshima et al., 1994
;
Sullivan et al., 2000
;
Vázquez-Martínez et al., 2003
). Consistent with spatiotemporal embryonic
dRyR
expression and the role of cytoplasmic calcium management (
Li et al., 2025
), we show that
dRyR
loss of function and RNAi knockdown in developing muscles cause severe developmental muscle defects. We found that
dRyR
promoted myogenic differentiation and was required for myotube growth associated with myoblast fusion and followed by myonuclear spreading within the myotubes. Our observations in
Drosophila
are consistent with the promoting role of
RYR1
in the calcium-dependent myoblast-to-myotube fusion process reported by the Avinoam lab in an in vitro myogenic differentiation system (
Eigler et al., 2021
). Interestingly, our analyses of embryonic muscle phenotypes of hypomorph
dRyR
16
embryo reveal that
dRyR
, in addition to its major promyogenic role, may also negatively influence myogenic differentiation. A negative influence on myogenic differentiation and, in particular, on myoblast fusion has recently been reported in mouse
RYR1
mutant myoblast culture (
Tourel et al., 2025
). It was suggested that this early developmental role of
RYR1
was calcium-independent.
dRyR
might thus play a dual role in myogenesis: (i) as a calcium-independent negative regulator of first myoblast-to-myoblast fusion events and (ii) as a positive regulator of myogenic differentiation acting in later steps of myogenesis in a calcium-dependent way to promote myoblast-to-myotube fusion and muscle fiber growth. This major pro-myogenic
dRyR
function is further supported by the
dRyR
gain-of-function phenotypes.
The knowledge gained on
dRyR
and its muscle-associated functions prompted us to use
Drosophila
to test the impact of one human
RYR1
mutation with unknown significance. We chose undiagnosed recessive
p.Met4881Ile
variant mutation identified in a patient with congenital myopathy phenotypes, severely impaired mobility, and accumulation of filamentous aggregates in muscle fibers. We found that
Drosophila
larvae carrying
p.Met4881Ile
mutation in the
dRyR
gene showed
dRyR RNAi
-like phenotypes with impaired larval mobility and significantly impaired sarcomeric muscle structure.
In all, we assessed eight different functional and structural muscle variables showing that
p.Met4881Ile
variant mutation consistently impaired larval muscle performance and changed muscle size and structure. This suggests that the
p.Met4881Ile
mutation impairs
dRyR
function and is likely pathogenic.
To conclude, we report functional analysis of
dRyR
, the sole fruit fly
RyR
gene, and show that in addition to ensuring contractile properties of differentiated striated muscle, it plays a key pro-myogenic role during muscle development. Our findings advocate
Drosophila
for modeling and testing the impact of human
RYR1
variant mutations of unknown significance.
Materials and methods
Key resources table
Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
Gene ( Drosophila melanogaster ) | RyR | FBgn0011286 | ||
Genetic reagent ( D. melanogaster ) | p.Met4881Ile RYR1 VUS | This paper | Generated by CRISPR-Cas9 homologous recombination genome editing Available on request | |
Genetic reagent ( D. melanogaster ) | C57-GAL4 | Bloomington Drosophila Stock Center | BDSC:32556; FLYB: FBti0016293; RRID: BDSC_32556 | GAL4 driver line FlyBase symbol: P{GawB}C57 |
Genetic reagent ( D. melanogaster ) | Hand-GAL4 | Laurent Perrin, TAGC, Marseille, France | GAL4 driver line | |
Genetic reagent ( D. melanogaster ) | Lms-GAL4 | Bloomington Drosophila Stock Center (unavailable) | BDSC:46861 | GAL4 driver line FlyBase symbol: P{GMR88F09-GAL4}attP2 |
Genetic reagent ( D. melanogaster ) | RyR TRIP | Bloomington Drosophila Stock Center | BDSC:29445; FLYB: FBti0129073; RRID: BDSC_29445 | RNAi line FlyBase symbol: P{TRiP.JF03381}attP2 |
Genetic reagent ( D. melanogaster ) | SERCA TRIP | Bloomington Drosophila Stock Center | BDSC_44581; FLYB: FBti0158759; RRID: BDSC_44581 | RNAi line FlyBase symbol: P{TRiP.HMS02878}attP2 |
Genetic reagent ( D. melanogaster ) | Cam TRIP | Bloomington Drosophila Stock Center | BDSC:34609; FLYB: FBti0140942; RRID: BDSC_34609 | RNAi line FlyBase symbol: P{TRiP.HMS01318}attP2 |
Genetic reagent ( D. melanogaster ) | mCherry RNAi | Bloomington Drosophila Stock Center | BDSC:35785; FLYB: FBti0143385; RRID: BDSC_35785 | RNAi line FlyBase symbol: P{VALIUM20-mCherry.RNAi}attP2 |
Genetic reagent ( D. melanogaster ) | UAS-GCaMP | Bloomington Drosophila Stock Center | BDSC:32236; FLYB: FBti0131954; RRID: BDSC_32236 | UAS line FlyBase symbol: P{20XUAS-GCaMP3}attP2 |
Genetic reagent ( D. melanogaster ) | UAS-RyR | Howard A Nash, University of Maryland College Park, Rockville, USA | UAS line | |
Genetic reagent ( D. melanogaster ) | UAS-RedStinger | Bloomington Drosophila Stock Center | BDSC:8547; FLYB: FBti0040830; RRID: BDSC_8547 | UAS line FlyBase symbol: P{UAS-RedStinger}6 |
Genetic reagent ( D. melanogaster ) | UAS-RedStinger | Bloomington Drosophila Stock Center | BDSC:8546; FLYB: FBti0040829; RRID: BDSC_8546 | UAS line FlyBase symbol: P{UAS-RedStinger}4 |
Genetic reagent ( D. melanogaster ) | UAS-LacZ | Bloomington Drosophila Stock Center | BDSC:1776; FLYB: FBti0002128 RRID: BDSC_1776 | UAS line FlyBase symbol: P{UAS-lacZ.B}Bg4-1-2 |
Genetic reagent ( D. melanogaster ) | UAS-lifeAct-GFP | Bloomington Drosophila Stock Center | BDSC:35544; FLYB: FBti0143326 RRID: BDSC_35544 | UAS line FlyBase symbol: P{UAS-Lifeact-GFP}VIE-260B |
Genetic reagent ( D. melanogaster ) | RyR 16 | Bloomington Drosophila Stock Center | BDSC:6812; FLYB: FBal0117664 RRID: BDSC_6812 | FlyBase symbol: RyR 16 |
Genetic reagent ( D. melanogaster ) | w 1118 | Bloomington Drosophila Stock Center | BDSC:3605; FLYB: FBal0117664; RRID: BDSC_3605 | FlyBase symbol: FBal0018186 |
Antibody | anti-dlg1 (Mouse monoclonal) | Developmental Studies Hybridoma Bank (DSHB) | Cat#: 4F3 | IF(1:50) |
Antibody | mouse anti-sls (Kettin) (Mouse monoclonal) | Developmental Studies Hybridoma Bank (DSHB) | Cat#: 1B8-3D9 | IF(1:50) |
Antibody | anti-GFP (Goat polyclonal) | Abcam | Cat#: ab5450 | IF(1:500) |
Antibody | anti-ATP5A (Mouse monoclonal) | Abcam | Cat#: ab14748 | IF(1:200) |
Antibody | anti-Actin (Rat monoclonal) | Abcam | Cat#: ab50591 | IF(1:500) |
Antibody | anti-beta galactosidase (Chicken polyclonal) | Abcam | Cat#: ab9361 | IF(1:1000) |
Recombinant DNA reagent | PCFD5 plasmid | Adgene | Plasmid #73914 | |
Sequence-based reagent | RyR F | This paper | PCR primers | 5’- TGCAGAGCAGCCGGAGGATGAC |
Sequence-based reagent | RyR R | This paper | PCR primers | 5’- ATCAGACGCGGCGAATCCGCAG |
Drosophila
strains and genetics
Request a detailed protocol
Fly stocks were maintained at 25 °C on standard fly food.
The targeted expression experiments were performed using the UAS-GAL4 system (
Brand and Perrimon, 1993
) on the following GAL4 and UAS lines: C57-GAL4 (Bl32556); Hand-GAL4 (kindly provided by L. Perrin; TAGC, Aix-Marseille University, France); UAS-dRyR RNAi (BL29445); Lms-GAL4 (BL46861); UAS-mCherry RNAi (BL35785); UAS-dRyR (kindly provided by H. Nash University of Maryland College Park, Rockville, USA), UAS-lifeAct-GFP (BL35544), UAS-dsRed NLS (BL8547; BL8546), UAS-LacZ (BL1776), UAS-Cam RNAi (Bl34609), UAS-SERCA RNAi (BL44581), and UAS-GCaMP3 (BL32236). The RyR
16
/CyO Wg LacZ (BL6812) was used as hypomorphic mutant and the
w
1118
strain was used as wild-type.
Phylogenetic analysis
Request a detailed protocol
Initial trees for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the maximum composite likelihood (MCL) approach and then selecting the topology with superior log likelihood value (
Kumar et al., 2018
). The tree is drawn to scale, with branch lengths measured in number of substitutions per site. This analysis involved seven nucleotide sequences. Codon positions included were first + second + third + Noncoding. There were 17,322 positions in the final dataset.
Immunohistochemistry
Request a detailed protocol
Antibody staining was performed using standard protocol. Embryos were fixed in 4% formaldehyde and blocked in NGS serum to remove non-specific epitopes. They were incubated overnight at 4 °C with primary antibodies followed by secondary antibodies for 2 hr at RT.
Third instar larvae were dissected and fixed in 4% paraformaldehyde for 20 min as previously described (
Lavergne et al., 2020
). The fly hearts were dissected (
Fink et al., 2009
;
Ocorr et al., 2007
) and fixed for 15 min in 4% formaldehyde and the immunostaining procedure was performed as described (
Auxerre-Plantié et al., 2019
).
The following primary antibodies were used in this study: guinea pig anti-dRYR antibody (1:200, kindly provided by Robert Scott and Benjamin White from NIH/NIMH Institute and previously described in
Gao et al., 2013
), mouse anti-Dlg (1–50, DSHB, 4F3), rat anti-actin (1–500, Abcam, ab 50591), mouse anti-kettin (1:50 DSHB 1B8-3D9), mouse anti-ATP5A (1:200 Abcam, ab 14748), goat anti-GFP (1:500, Abcam, ab 5450), and chicken anti-β galactosidase (1:1000, Abcam, ab 9361). Rhodamine phalloidin (Thermo Fischer Scientific) was used to reveal actin filaments in the heart and muscles. Fluorescent secondary antibodies (Jackson ImmunoResearch) were used to detect primary antibodies.
Muscle characteristics measurements
Request a detailed protocol
All analyses of muscle length and sarcomere size were performed on fixed larval muscle preparations in a relaxed state. Acquired confocal images were analysed in Fiji using the
line tool. Analyze – Measure
tool was then applied to obtain muscle length values and measurements were analysed with Prism. Sarcomere size and number were calculated using
Analyze – Plot profile
Fiji tool. The sarcomere size was measured between peaks corresponding to Z-disc (revealed with Z-line specific marker) on approximately 100 μm of muscle length. Sarcomere measurements were then analysed with Prism.
DAPI-stained nuclei were counted in Z-stacks of confocal views of VL3 larval muscle and data analyzed with Prism. About 30 larval muscles from 6 to 8 larval filets were analysed for each measurement.
In situ hybridization chain reaction – HCR
Request a detailed protocol
In this study, we used the two-stage in situ HCR protocol described by
Choi et al., 2016
. This technique detects and amplifies specific transcripts by the direct binding of probes to nucleic target sequences without additional long-lasting enzymatic reaction. We planned four different mRNA probes targeting alternative exons numbered 10, 11, 22, and 23.
We used a Molecular Instruments HCR kit containing a DNA probe set, a DNA HCR amplifier B1-Alexa fluor 488, B2-fluor 532 and hybridization, wash and amplification buffers.
Fixed samples were pre-hybridized at 65 °C for 2 hr followed by hybridization steps overnight at 45 °C. After several washes, the amplification step was performed overnight at RT in the dark. We used TRJ244 HCR amplifier B1, RTJ245 HCR amplifier B2, RTJ256 HCR amplifier B2, RTJ 247 HCR amplifier B1. Target mRNA sequence information remains at the discretion of Molecular Instruments Company. Excess of hairpins was removed by several washes with 5XSSCT solution at room temperature.
Imaging
Request a detailed protocol
Samples were mounted in Vectashield with DAPI (Vector Laboratories, Inc Burlingame, CA) and a Leica SP8 confocal microscope was used for image acquisition and for time-lapse imaging of living embryos. In vivo imaging of lateral muscle was performed from late stage 14 to stage 16 every 3 min. We used Adobe Photoshop and ImageJ for image processing.
Genome editing
Request a detailed protocol
To generate the
Drosophila
model of human
RYR1
VUS p.Met4881Ile we applied the CRISPR-Cas9 homologous recombination genome editing approach with the use of single-strand oligo donor (ssODN) and gRNA. A 20nt 5’-
AAACGCTTCGTGTTCCATCTGTAC
-3’ gRNA targeting the
dRYR
region close to the mutation site was designed using the OPTIMAL Target Finder platform (
Gratz et al., 2014
) and cloned into PCFD5 plasmid (Adgene). Single-strand ssODN donor of 115 nt in size with sequence modification TTG to ATC was generated by IDT Company. Both components were injected at a concentration of 100 ng/μl by Best Gene into nos-Cas 9 (III-attp2) flies. PCR molecular screening with a pair of primers: Forward 5’-
TGCAGAGCAGCCGGAGGATGAC
; Reverse 5’-
ATCAGACGCGGCGAATCCGCAG
and Sanger sequencing were used to identify progenies carrying the edited sequence. Genetic crosses established homozygous
Drosophila
lines carrying the RyR1 p.Met4881Ile variant mutation.
Functional tests of muscle performance
Request a detailed protocol
Motility tests were carried out on 30 third instar larvae as previously described (
Picchio et al., 2013
). The motility test was performed by recording the number of peristaltic contractions executed by the larva in 1 min on a grape medium plate. The righting test consisted in placing the larva on its back and measuring the time it took to revert to its crawling position. For the locomotor test, a track 2 mm wide, 5 mm deep and 5 cm long was prepared on a grape medium plate. Larvae were placed on the test track and the time they took to crawl a distance of 5 cm was recorded.
Heartbeat analyses
Request a detailed protocol
The cardiac activity analyses of adult
Drosophila
hearts were performed on 1-week-old female flies using the Semi-automated Optical Heartbeat Analysis (SOHA) approach protocol of
Fink et al., 2009
. For each experiment, about 20 flies were analyzed. The flies were anesthetized with Fly Nap, dissected in an oxygenated, artificial hemolymph composed of 108 mM NaCl, 5 mM KCl, 2 mM CaCl
2
, 8 mM MgCl
2
, 1 mM NaH
2
PO
4
, 4 mM NaHCO
3
, 10 mM sucrose, 5 mM trehalose, 5 mM HEPES (pH 7.1). The beating hearts were filmed by digital camera on 30 s movie with the speed of 150 frames/s (Digital camera C9300, Hamamatsu, McBain Instruments, Chatsworth, CA). The SOHA program, based on Matlab R2009b software, was used for film analysis (
Fink et al., 2009
).
Mitochondria area quantifications
Request a detailed protocol
The total area of mitochondria was identified by Otsu thresholding on Fiji for internal muscle z-planes. The quantifications for z-planes were averaged for each of 25–30 different VL3 muscles according to
Zhang et al., 2024
.
Statistic
Request a detailed protocol
All statistical analyses were performed using Prism (v9.5.1, GraphPad Software, La Jolla, CA, USA). The
t
-test was used to compare control to variant context and one-way ANOVA tests were used for comparisons with more than two datasets. Bar plot represents the mean and the standard deviation. On the figures, statistical comparisons of sample vs control are indicated as ****
p
≤0.0001; ***
p
≤0.001; **
p
≤0.01; *
p
≤0.05; ns>0.05.
Data availability
All behavioral, phenotypical and image analyses datasets that were used to generate graphs in Figures 1–6 are available as source data files.
References
Straightjacket/α2δ3
deregulation is associated with cardiac conduction defects in myotonic dystrophy type 1
eLife
8
:e51114.
PubMed
Google Scholar
Gelsolin and dCryAB act downstream of muscle identity genes and contribute to preventing muscle splitting and branching in Drosophila
Scientific Reports
11
:13197.
PubMed
Google Scholar
As time flies by: Investigating cardiac aging in the short-lived Drosophila model
Biochimica et Biophysica Acta. Molecular Basis of Disease
1865
:1831–1844.
PubMed
Google Scholar
Targeted gene expression as a means of altering cell fates and generating dominant phenotypes
Development
118
:401–415.
PubMed
Google Scholar
Mapping a multiplexed zoo of mRNA expression
Development
143
:3632–3637.
PubMed
Google Scholar
Alternative splicing and insect ryanodine receptor
Archives of Insect Biochemistry and Physiology
102
:e21590.
PubMed
Google Scholar
Evolution of vertebrate ryanodine receptors family in relation to functional divergence and conservation
International Heart Journal
58
:969–977.
PubMed
Google Scholar
ERK1/2 inhibition promotes robust myotube growth via CaMKII activation resulting in myoblast-to-myotube fusion
Developmental Cell
56
:3349–3363.
PubMed
Google Scholar
Quantitative RyR1 reduction and loss of calcium sensitivity of RyR1Q1970fsX16+A4329D cause cores and loss of muscle strength
Human Molecular Genetics
28
:2987–2999.
PubMed
Google Scholar
Ryanodine receptor calcium release channels
Physiological Reviews
82
:893–922.
PubMed
Google Scholar
A new method for detection and quantification of heartbeat parameters in Drosophila, zebrafish, and embryonic mouse hearts
BioTechniques
46
:101–113.
PubMed
Google Scholar
Regulation of the RYR1 and RYR2 Ca2+ release channel isoforms by Ca2+-insensitive mutants of calmodulin
Biochemistry
42
:2740–2747.
PubMed
Google Scholar
Drosophila ryanodine receptors mediate general anesthesia by halothane
Anesthesiology
118
:587–601.
PubMed
Google Scholar
Alternative splicing of ryanodine receptors modulates cardiomyocyte Ca2+ signaling and susceptibility to apoptosis
Circulation Research
100
:874–883.
PubMed
Google Scholar
Highly specific and efficient CRISPR/Cas9-catalyzed homology-directed repair in Drosophila
Genetics
196
:961–971.
PubMed
Google Scholar
Structural insight into ryanodine receptor channelopathies
Frontiers in Pharmacology
13
:897494.
PubMed
Google Scholar
Drosophila homologs of two mammalian intracellular Ca(2+)-release channels: identification and expression patterns of the inositol 1,4,5-triphosphate and the ryanodine receptor genes
Development
116
:967–975.
PubMed
Google Scholar
The zebrafish reference genome sequence and its relationship to the human genome
Nature
496
:498–503.
PubMed
Google Scholar
The pharmacology of ryanodine
Pharmacological Reviews
21
:1–25.
PubMed
Google Scholar
A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences
Journal of Molecular Evolution
16
:111–120.
PubMed
Google Scholar
Intracellular calcium mobilization is required for sonic hedgehog signaling
Developmental Cell
45
:512–525.
PubMed
Google Scholar
MEGA X: molecular evolutionary genetics analysis across computing platforms
Molecular Biology and Evolution
35
:1547–1549.
PubMed
Google Scholar
Ryanodine receptors: structure, expression, molecular details, and function in calcium release
Cold Spring Harbor Perspectives in Biology
2
:a003996.
PubMed
Google Scholar
Ryanodine receptor physiology and its role in disease
Advances in Experimental Medicine and Biology
740
:217–234.
PubMed
Google Scholar
Drosophila
adult muscle precursor cells contribute to motor axon pathfinding and proper innervation of embryonic muscles
Development
147
:dev183004.
PubMed
Google Scholar
Regulation of calcium homeostasis in endoplasmic reticulum-mitochondria crosstalk: implications for skeletal muscle atrophy
Cell Communication and Signaling
23
:17.
PubMed
Google Scholar
Presynaptic ryanodine receptors are required for normal quantal size at the
Caenorhabditis elegans
neuromuscular junction
The Journal of Neuroscience
25
:6745–6754.
PubMed
Google Scholar
Book
Ryanodine receptor calcium release channels: an evolutionary perspective
In: Islam S, editors.
Calcium Signaling
. Netherlands: Springer. pp. 159–182.
Google Scholar
Targeting ryanodine receptors to treat human diseases
The Journal of Clinical Investigation
133
:e162891.
PubMed
Google Scholar
unc-68 encodes a ryanodine receptor involved in regulating
C. elegans
body-wall muscle contraction
The Journal of Cell Biology
134
:885–893.
PubMed
Google Scholar
A comparative study indicates both positive AND purifying selection within ryanodine receptor (RyR) genes, as well as correlated evolution
Journal of Experimental Zoology. Part A, Ecological Genetics and Physiology
321
:151–163.
PubMed
Google Scholar
Two ryanodine receptor isoforms in nonmammalian vertebrate skeletal muscle: possible roles in excitation-contraction coupling and other processes
Progress in Biophysics and Molecular Biology
105
:134–144.
PubMed
Google Scholar
Drosophila as a model to study cardiac aging
Experimental Gerontology
46
:326–330.
PubMed
Google Scholar
KCNQ potassium channel mutations cause cardiac arrhythmias in Drosophila that mimic the effects of aging
PNAS
104
:3943–3948.
PubMed
Google Scholar
Alpha and beta isoforms of ryanodine receptor from chicken skeletal muscle are the homologues of mammalian RyR1 and RyR3
The Biochemical Journal
315
:207–216.
PubMed
Google Scholar
Novel Drosophila model of myotonic dystrophy type 1: phenotypic characterization and genome-wide view of altered gene expression
Human Molecular Genetics
22
:2795–2810.
PubMed
Google Scholar
Vertebrate genome evolution and the zebrafish gene map
Nature Genetics
18
:345–349.
PubMed
Google Scholar
Characterization of the ryanodine receptor/channel of invertebrate muscle
American Journal of Physiology-Regulatory, Integrative and Comparative Physiology
274
:R494–R502.
Google Scholar
Ca
2+
as a coordinator of skeletal muscle differentiation, fusion and contraction
The FEBS Journal
289
:6531–6542.
PubMed
Google Scholar
Molecular evolutionary genetics analysis (MEGA) for macOS
Molecular Biology and Evolution
37
:1237–1239.
PubMed
Google Scholar
Reduced SERCA function preferentially affects wnt signaling by retaining e-cadherin in the endoplasmic reticulum
Cell Reports
26
:322–329.
PubMed
Google Scholar
The ryanodine receptor is essential for larval development in
Drosophila melanogaster
PNAS
97
:5942–5947.
Google Scholar
Isolation and characterization of a gene for a ryanodine receptor/calcium release channel in
Drosophila melanogaster
FEBS Letters
337
:81–87.
https://doi.org/10.1016/0014-5793(94)80634-9
PubMed
Google Scholar
RyR1 is involved in the control of myogenesis
Cells
14
:158.
PubMed
Google Scholar
Biochemical characterization, distribution and phylogenetic analysis of
Drosophila melanogaster
ryanodine and IP3 receptors, and thapsigargin-sensitive Ca2+ ATPase
Journal of Cell Science
116
:2483–2494.
PubMed
Google Scholar
Molecular characterization of a ryanodine receptor gene in the rice leaffolder, cnaphalocrocis medinalis (Guenée)
PLOS ONE
7
:e36623.
Google Scholar
Ryanodine receptors, a family of intracellular calcium ion channels, are expressed throughout early vertebrate development
BMC Research Notes
4
:541.
PubMed
Google Scholar
Molecular cloning of cDNA encoding a drosophila ryanodine receptor and functional studies of the carboxyl-terminal calcium release channel
Biophysical Journal
78
:1270–1281.
https://doi.org/10.1016/S0006-3495(00)76683-5
PubMed
Google Scholar
Mechanoresponsive regulation of myogenesis by the force-sensing transcriptional regulator Tono
Current Biology
34
:4143–4159.
PubMed
Google Scholar
Article and author information
Author details
Monika Zmojdzian

"This ORCID iD identifies the author of this article:"
0000-0001-6174-2719
Funding
Institut National de la Santé et de la Recherche Médicale (RYR-ClassifAI)
John Rendu
Krzysztof Jagla
Department of Agriculture, Food and the Marine, Ireland (MyoNeuralp)
Monika Zmojdzian
Agence Nationale de la Recherche (iSITE CAP2025)
Krzysztof Jagla
Catherine Sarret
Bekker Programme (BPN/BEK/2021/2/00006)
Magda Dubinska-Magiera
Marta Migocka-Patrzałek
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Drosophila Bloomington Stock Centers for Drosophila lines and L Mouty for the technical assistance. This study was supported by the 'Priority Research Programme on Rare Diseases' of the French Investments for the Future Programme, by the AFM-Telethon strategic grant to MyoNeurAlp consortium, the Reference Centre for Neuromuscular Diseases grant to MZ and CS, ANR-iSITE-2025 grant to MZ, by the French Government Scholarship grant to MMP and The Polish National Agency for Academic Exchange within The Bekker Programme (grant no. BPN/BEK/2021/2/00006) to MDM.
Ethics
This study was conducted in accordance with the ethical standards of the institutional and national research committees. Ethical approval was obtained from the appropriate institutional ethics committee under the reference 38RC21.0399.
Version history
Cite all versions
You can cite all versions using the DOI
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2026, Zmojdzian, Jagla et al.
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
241
views
7
downloads
0
citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads
(link to download the article as PDF)
Article PDF
Figures PDF
Open citations
(links to open the citations from this article in various online reference manager services)
Mendeley
Cite this article
(links to download the citations from this article in formats compatible with various reference manager tools)
Drosophila
ryanodine receptor gene triggers functional and developmental muscle properties and could be used to assess the impact of human
RYR1
mutations
eLife
15
:RP111053.





