 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库The NTR/Prodrug Revolution: Tools for Controlling Cell Loss and Regeneration
Abstract
Here, we review the history, advancements, and broad utility of the NTR/prodrug system, and suggest future strategies for developing versatile ablation models. As a chemogenetic tool, the nitroreductase (NTR)/prodrug system enables precise spatiotemporal control over cell ablation. The technology leverages bacterial nitroreductase enzymes (e.g.,
nfsB
) to convert inert prodrugs into cytotoxic agents, thereby allowing researchers to induce targeted cell death. Although the NTR/prodrug approach was first implemented in transgenic mice, it was subsequently adapted to zebrafish, where it has been extensively optimized and applied. Consequently, zebrafish remain the primary focus of this review. Nevertheless, the utility of the NTR/prodrug system has expanded to other important model organisms, including
Drosophila, Nematostella
,
Xenopus
, medaka, and rats, enabling detailed studies of tissue damage and regeneration. This review highlights how the NTR system has been deployed to model a spectrum of human diseases, including Parkinson’s disease, retinal degeneration, demyelinating disorders, and kidney disease. These models provide valuable platforms to study pathogenesis
in vivo
. Furthermore, the precise and controllable nature of NTR ablation makes it an ideal tool for high-throughput chemical and genetic screens aimed at discovering pro-regenerative and protective compounds. The development of NTR2.0, an enzyme variant with over 100-fold greater activity, along with more potent prodrugs such as ronidazole (RNZ), has dramatically broadened experimental possibilities. These improvements permit chronic ablation and long-term disease modeling at well-tolerated drug concentrations. Here we present some key considerations including transgenic design for optimal cell-type specificity, calibrating expression levels for desired ablation kinetics, and suitable controls to allow interpretation. These best practices will allow the researcher to develop a precise, reproducible, and versatile platform for either modeling human disease or dissecting regenerative mechanisms.
Introduction
Why Regeneration?
We often regard ourselves as having limited regenerative capacity, yet numerous human tissues exhibit substantial renewal: the liver restores lost mass, the skin undergoes continuous stem cell-driven turnover, skeletal muscle repairs through satellite cells, bone heals via remodeling, blood is replenished by hematopoietic stem cells, the intestinal epithelium renews within days, and the endometrium regenerates cyclically. However, the regenerative capacity of human tissues is modest when contrasted with that of many other species, in which entire organs or appendages can be restored following injury [
1
,
2
]. This capacity is profound in many invertebrates; for instance, hydra and planarians can regenerate complete organisms from minute fragments [
3
].
Understanding these enviable examples provides a framework for uncovering the cellular and molecular mechanisms that organisms employ to repair or replace damaged tissues, with the ultimate goal of developing new strategies to combat human injuries, degenerative diseases, and age-related decline [
4
,
5
]. An essential requirement for this research is the ability to induce controlled, reproducible injury to study the subsequent repair processes.
The laboratory mouse
(Mus musculus)
is a critical vertebrate model for human biology and has been essential for developing inducible, cell-type-specific genetic tools in mammals. Like adult humans, adult mice exhibit regenerative and reparative abilities across several tissues; however, their capacity for epimorphic regeneration (regrowth of complex structures such as heart tissue or appendages) is largely confined to early postnatal stages. For developing precise ablation tools, a genetically tractable and cost-effective vertebrate with robust adult regeneration is desirable. The zebrafish (
Danio rerio)
is uniquely suited for this role. Its external development, transparency, and high fecundity facilitate direct
in vivo
visualization and high-throughput screening [
6
]. Critically, zebrafish exhibit widespread regenerative abilities in adulthood, fully restoring complex organs including the heart [
7
,
8
], retina [
9
,
10
], spinal cord [
11
,
12
], and fins [
13
,
14
]. These attributes make zebrafish a practical system for dissecting the mechanisms of vertebrate regeneration [
15
].
Methods of Ablation in Zebrafish
Regeneration studies rely on experimental injuries, and the nature of the injury profoundly shapes the ensuing repair response. In zebrafish, the major injury paradigms can be broadly grouped into the following categories:
1. Physical Injury
: Direct tissue damage via surgical intervention that have been used successfully to study adult regeneration, including fin amputation [
16
], ventricular resection [
7
], brain lesioning [
17
], scale removal [
18
,
19
] and transection of both tendons [
20
] and ligaments [
21
]. Severing of the spinal cord predictably leads to paralysis in zebrafish, but as testament to their regenerative capacity, full mobility is returned by 8 weeks [
11
,
22
,
23
].
Several non-surgical physical methods can be used to induce tissue damage in animal models, with their suitability largely determined by the subject’s size and developmental stage. In adult models, approaches include cryoinjury to simulate myocardial infarction [
24
,
25
], intense light exposure to trigger photoreceptor degeneration in retinal regeneration studies [
26
], and acoustic trauma to model inner-ear damage and hearing loss recovery [
27
,
28
]. The acoustic method has also been applied to larval zebrafish to target lateral line hair cells [
29
]. Larval and embryonic zebrafish, owing to their small size and optical transparency, permit highly precise laser ablation of single neurons [
30
–
32
] or even individual cardiomyocytes [
33
]. Additionally, thermal injury has been employed in larvae to model burns and skin regeneration, revealing rapid inflammatory cell recruitment [
34
] and keratinocyte migration, but impaired sensory axon regeneration compared to mechanical injury [
35
].
Regardless of the method, each of these physical injury approaches perturbs tissue integrity in characteristic ways, disrupting multiple components of tissue architecture, including extracellular matrix, mesenchymal organization, and local vasculature. This disruption drives rapid physiological and robust inflammatory responses involving neutrophils and macrophages, all of which critically influence regenerative outcomes [
34
–
36
].
2. Cell-Specific Toxins
: While physical injury has provided key insights into regenerative biology, these approaches are often constrained by limited precision and cell-type specificity. Pharmacological methods offer a complementary strategy, using small molecules to induce targeted, dose-controlled damage.
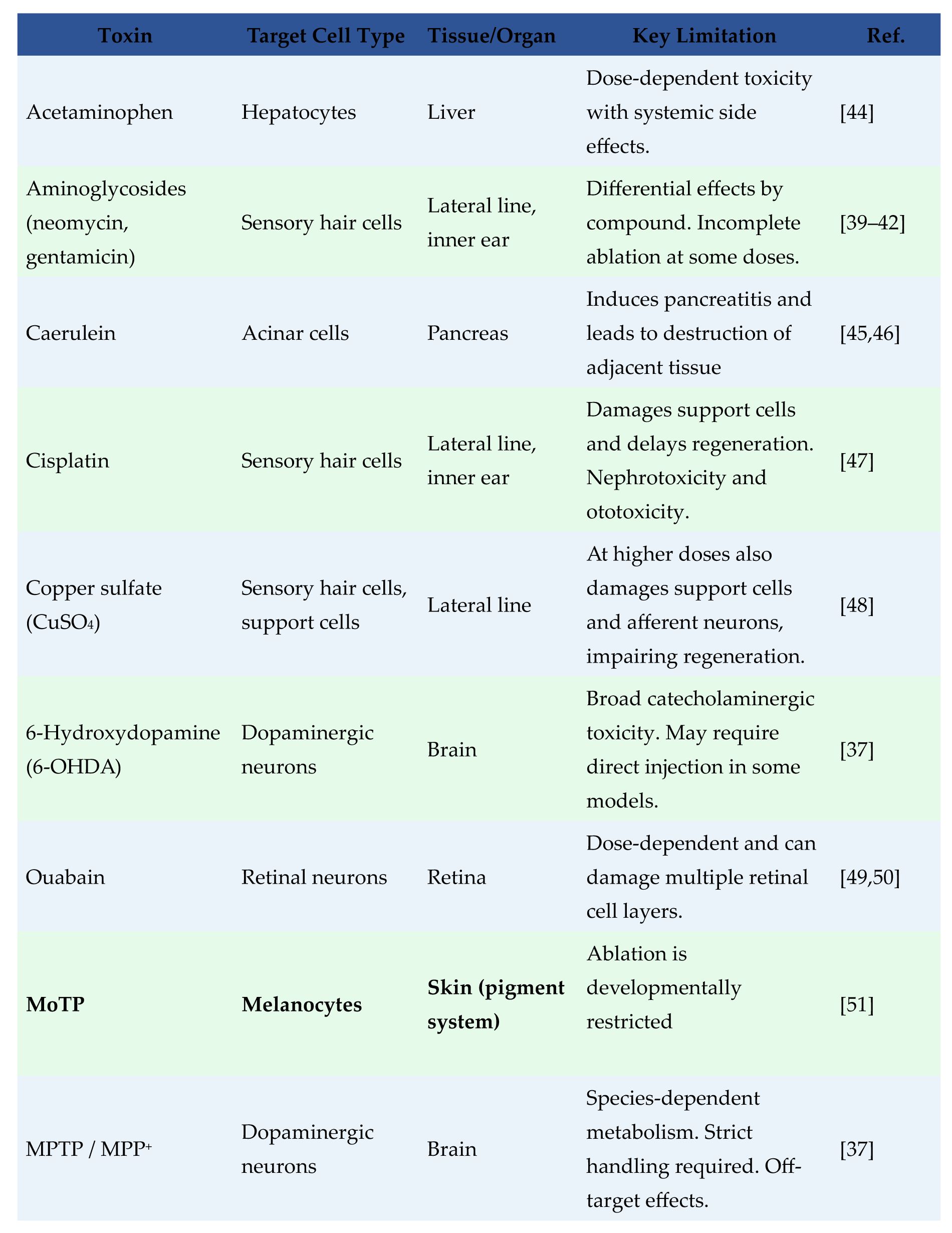
Table 1
summarizes representative compounds used for cell-specific ablation in zebrafish. However, many of these agents introduce off-target effects, ranging from dose-dependent systemic toxicity to unintended tissue injury, which can make downstream regenerative responses harder to interpret. For example, MPTP and MPP
+
mainly target dopaminergic neurons but can also impact noradrenergic and serotonergic systems via uptake through their neurotransmitter transporters [
37
,
38
]. Similarly, aminoglycoside antibiotics, while commonly used to ablate lateral line hair cells, can also damage support cells and afferent neurons at higher concentrations, and exhibit compound-specific differences in ototoxicity [
39
–
42
]. Streptozotocin, widely used to ablate pancreatic β-cells, is known to cause broader cytotoxicity beyond the target population [
43
]. To achieve higher precision and reduce off-target effects as well as expand the kinds of cell that can be ablated, the field has increasingly turned to genetic strategies. These methods allow damage to be targeted with exquisite specificity to predefined cell types.
3. Optogenetic cell ablation
: Due to the transparent nature of young zebrafish, this model is highly amenable to optogenetic techniques that enable precise, non-invasive control of cellular signaling, neuronal circuit activity, and targeted cell ablation (see reviews [
52
,
53
]). KillerRed is a genetically encoded photosensitizer that produces reactive oxygen species (ROS) upon green or yellow light illumination, killing nearby cells [
54
,
55
]. When expressed under cell-specific promoters, this technique allows researchers to eliminate defined cell populations in living embryos or larvae with precise spatiotemporal control [
54
–
56
]. A key advantage of this method is its speed, as KillerRed-expressing cells can be ablated within hours of light exposure. This approach is particularly useful for modeling diseases in which ROS-mediated cell death is central, such as neurodegeneration [
57
] and cardiomyopathies [
58
], but it has not yet been applied in conventional regeneration studies. Although speculative, this method appears best suited for discrete, optically accessible cells [
54
] and is likely to be impractical for ablation of large populations, whole tissues, or cells deep within the adult body, where localized illumination is more difficult. Because this method relies on controlled optical delivery via microscopy, it also limits throughput making high-volume studies challenging.
4. Chemogenetic, cell-specific ablation
: Chemogenetic ablation relies on transgene-driven, cell-specific expression of an exogenous protein that converts an otherwise innocuous chemical into a cytotoxic agent. By eliminating only the intended population and leaving neighboring tissues intact, these methods generate narrowly focused injuries whose regenerative responses differ substantially from those induced by broader, multi-tissue physical damage. Four such chemogenetic systems established in zebrafish include:
i. Human Diphtheria Toxin Receptor and Diphtheria toxin
: The human pathogen
Corynebacterium diphtheriae
produces diphtheria toxin (DT), which enters cells by binding the human diphtheria toxin receptor (hDTR) [
59
,
60
]. Because endogenous receptors in non-primates do not bind DT efficiently, these animals are naturally resistant to the toxin. This strategy allows researchers to engineer specific cell types to express hDTR, making them uniquely susceptible to DT-induced ablation [
61
]. In mouse research, the DTR/DT system has become one of the most commonly employed strategies for conditional cell ablation, owing to its rapid and reliable elimination of targeted cell populations across many tissues. However, the method has caveats, like the fact that DT alone can cause kidney damage [
62
] and that repeated DT exposure induces production of neutralizing antibodies, preventing long-term or chronic ablation.
Although DTR/DT approaches are widely used in mouse studies, their application in zebrafish has been more limited [
63
,
64
]. A noteworthy example is the transgenic line generated by Jimenez et al., in which hDTR driven by the hair-cell-specific myo6b promoter enabled selective depletion of sensory hair cells following DT administration, with full regeneration occurring within days [
63
]. Some studies in zebrafish have instead expressed diphtheria toxin subunit A (DTA) directly in target cells [
65
,
66
], although constitutive DTA expression lacks temporal control and is not suitable for regeneration studies. Temporal control can be introduced through Cre/lox-inducible DTA systems [
67
] but issues such as recombination efficiency, mosaicism, and promoter leakiness may limit precision [
68
].
ii. Inducible caspase systems
: Caspase cascade in apoptosis begins with upstream initiator caspases (e.g., caspase 8) activated by dimerization, which then activates effector caspases (e.g., caspase 3) to execute cellular dismantling [
69
–
71
]. In zebrafish, researchers have exploited induction of caspase-8 dimerization to achieve temporal and spatial control of cell-specific ablation in two main ways: 1) activated by the FK1012 chemical inducer of dimerization [
64
,
72
]; and, 2) expression of a fusion between caspase and the modified estrogen receptor ligand-binding domain (ERT2), whose activity is induced by binding of tamoxifen [
73
–
75
]. Compared to FK1012, tamoxifen pharmacokinetics are better characterized in zebrafish, and a convincing study showing lifelong regeneration of cerebellar Purkinje cells in zebrafish strongly supports the use of the ER-T2/tamoxifen inducible ablation approach for probing cell loss and recovery [
73
].
iii. HSV-TK
: The herpes simplex virus thymidine kinase (HSV-tk) has been applied in zebrafish for conditional cell ablation. Transgenic expression of this ‘suicide gene’ (here defined as a gene encoding an enzyme that converts a nontoxic prodrug into a cytotoxin [
76
]) in a defined cell population converts the antiviral prodrug ganciclovir into toxic nucleotides that are lethal to proliferating cells, but this dependence on cell division has limited the utility of the approach [
77
].
iv. Nitroreductase (NTR)
: Bacterial nitroreductase genes (nfsB) are another class of suicide genes. These genes encode nitroreductases (NTR), enzymes that convert nitro-containing prodrugs (e.g., nitroimidazoles and nitrofurans) into cytotoxic metabolites. As a result, transgenic animal cells expressing NTR become vulnerable to treatment with prodrugs such as metronidazole (MTZ), ronidazole (RNZ), nifurpirinol (NFP) and CB1954. Unlike HSV-TK, the NTR/MTZ system operates as a ‘cell-cycle-independent’ method for targeted cell ablation and improved variants of NTR can efficiently ablate fully differentiated cell types [
78
,
79
].

The NTR/prodrug system is now a staple of zebrafish research, although, somewhat surprisingly, the approach was first developed in the mouse. Since then, it has been adapted for use in a wide range of model organisms, including:
Drosophila melanogaster
— Teeters
et al.
used NTR and RNZ to ablate multiple, diverse cell types during development, demonstrating rapid, temperature-independent ablation using a simple drug-feeding protocol [
80
].
Nematostella
— Gavgani
et al.
used NTR and NFP to ablate neurons and reveal their requirement in body-axis regeneration [
81
].
Xenopus laevis
— Two NTR/MTZ models, one targeting oligodendrocytes and the other rod photoreceptors, achieved reliable, cell-specific ablation with subsequent regeneration. One targeting study noted temperature-dependent NTR activity, with reduced efficiency below 22 °C [
82
–
85
].
Medaka
— Willems
et al.
used NTR and MTZ to conditionally ablate osteoblasts and assess regeneration following drug withdrawal [
86
].
Rat
— Kwak
et al.
applied NTR and CB1954 to ablate neonatal cerebellar and ventricular progenitors, resulting in ataxia and reduced cerebellar volume [
87
].
Zebrafish
—
The most extensively used model for NTR/prodrug-mediated cell ablation, with applications across a wide range of tissues to study regeneration and to model human disease
(
Figure 1
and
Supplemental Table 1
).
Mouse
— The first
in vivo
NTR/CB1954 transgenic models were generated in mouse, where NTR expression enabled selective ablation of T cells, and mammary luminal epithelial cells [
88
,
89
]. These initial studies were followed by ablation studies of adipocytes, neurons, and kidney podocytes, showing the versatility across diverse tissues of this ablation method [
90
–
92
]. However, most of these efforts were carried out in the context of evaluating NTR as a suicide-gene strategy for cancer therapy in humans, rather than a tool for studying cell regeneration. Subsequent work in mammals shifted toward the DTR system, which is the predominant method of chemogenetic, cell-specific ablation in mouse.
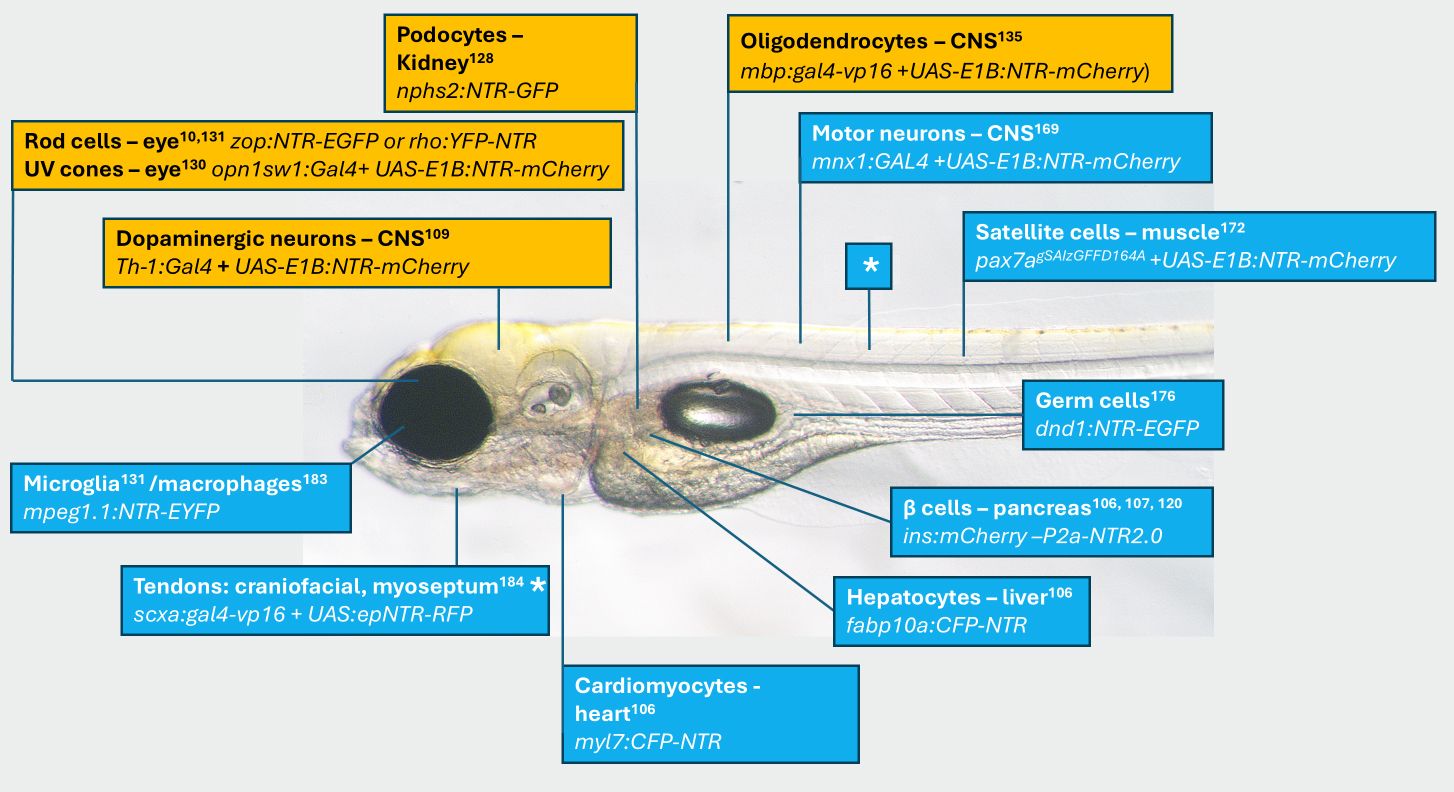
Commonly targeted cell types for ablation studies in zebrafish.
Supplemental Table 1
for more complete list. Image of a 5 days post-fertilization (dpf) casper zebrafish larva
182
with approximate position of cell type ablated. Name of cell, and transgene provided along with reference. Orange highlights indicate transgenic models used to study human pathologies. * same line two different tissues.
Development of Nitroreductase as a Suicide Gene
Bacterial nitroreductases (NTRs) from
E. coli (nfsA
and
nfsB
) were first charaderized in the 1970s-80s for their role in reducing nitroaromatics [
93
–
95
], a function later harnessed in the 1990s for Directed Enzyme Prodrug Therapy (DEPT) [
89
,
96
,
97
]. DEPT strategies use viral or antibody-based systems to deliver a ‘suicide gene’ product to tumors (e.g. NTR) [
98
]. Once localized, the enzyme converts an administered prodrug into a cytotoxic agent, selectively killing the cancer cells.
The prodrug CB1954 is harmless to human cells but becomes a powerful, DNA-damaging toxin after activation by NTR [
99
,
100
] Even though the NTR/CB1954 approach has been tested in clinical trials [
101
,
102
], its effectiveness in mitigating cancers is limited due to low NTR activity and slow prodrug metabolism. Despite these therapeutic shortcomings, this work established NTR/CB1954 as a potent conditional cell-killing strategy. However, this system is subject to a “bystander effect,” where activated metabolites diffuse into and kill neighboring cells. This attribute reduces the precision of targeted ablation and introduces ambiguity when assessing subsequent regeneration.
To overcome these limitations, researchers turned to alternative prodrugs. Metronidazole (MTZ), a nitroimidazole antibiotic, emerged as a particularly effective option as it is non-toxic to eukaryotic cells until reduced by bacterial NTR. Unlike CB1954, the activated metabolites of MTZ are short-lived and largely confined within the target cell, minimizing bystander effects [
103
–
105
]. This property made the NTR/MTZ system especially well-suited for regeneration and developmental studies, which benefit from precise, cell-specific ablation. In zebrafish, NTR/MTZ-mediated ablation consistently induces apoptosis across multiple tissues, as first shown in hepatocytes, cardiomyocytes, and pancreatic β-cells [
106
,
107
]. Across tissues, NTR/MTZ ablation elicits stress-linked apoptotic pathways, with hepatocytes showing elevated ROS and dopaminergic neurons exhibiting early mitochondrial impairment, both culminating in apoptosis [
108
,
109
]. The exact
coup de grâce
will likely vary by cell type, reflecting differences in metabolic activity, mitochondrial content, and intrinsic sensitivity to oxidative or genotoxic stress. Ablation kinetics are similarly context-dependent: NTR expression levels, cell identity, developmental stage, and MTZ dose all influence how rapidly and completely cells are eliminated. Defining how these variables shape NTR/MTZ-induced cytotoxicity across tissues remains an important direction for future work.
NTR/MTZ in Regenerative Studies
The NTR/MTZ system for cell-specific ablation was first described in two 2007 studies. Pisharath et al. placed the
E. coli
nfsB gene (NTR) under the zebrafish insulin promoter to express an NTR-mCherry fusion in pancreatic β cells; treatment with 10 mM MTZ produced complete β-cell loss without affecting neighboring α cells or exocrine tissue [
107
]. Curado et al. used cell-specific promoters to express CFP-NTR in cardiomyocytes, hepatocytes, and β cells, likewise demonstrating that MTZ induced highly targeted cell death with no detectable bystander effects [
106
]. Of note, NTR functions robustly whether fused to the N or C terminus of a fluorescent reporter, facilitating flexible transgene design [
104
,
110
]. In both studies, tissues regenerated after MTZ withdrawal, establishing the NTR/MTZ approach as a versatile, specific, inducible, and reversible tool for regeneration studies.
Unlike acute physical injury, NTR-mediated ablation induces apoptotic cell death and predominantly recruits macrophages rather than neutrophils [
108
]. This is an important consideration, as the type of cell death shapes the regenerative response. In zebrafish, macrophages are not merely phagocytic responders but essential regulators of regeneration: they clear apoptotic debris, modulate the inflammatory milieu, and release factors that support appropriate tissue remodeling [
111
,
112
].
This controlled context of NTR/MTZ ablation provides a platform for identifying the cells responsible for regeneration, a goal that can be achieved by pairing ablation with lineage tracing. For example, following targeted ablation of pancreatic β-cells, researchers used Cre-based lineage tracing to track the origins of regenerated endocrine cells. These studies revealed that Notch-responsive ductal cells called centroacinar cells (CACs) act as facultative progenitors that delaminate from the ducts, and replenish lost β-cells [
113
–
115
]. A parallel strategy applied to hepatocyte ablation uncovered an analogous regenerative mechanism in the liver where Notch-responsive biliary epithelial cells (BECs/cholangiocytes) delaminate, dedifferentiate, proliferate, and redifferentiate into new hepatocytes [
116
,
117
]. Combining single-cell RNA sequencing (scRNA-seq) of pancreatic ducts and hepatic ducts after β-cell/hepatocyte ablation has been used to map molecular mechanisms and identify intermediate progenitor states as new β-cells/hepatocytes are formed [
118
,
119
]. This integrated paradigm of targeted ablation, cre-based lineage tracing, and scRNA-seq provides a systematic framework for dissecting the mechanisms that drive tissue regeneration.
Combining Cre-based lineage tracing with NTR ablation can reveal the origins of regenerated cells. Another way to identify progenitors is to induce regeneration and then use NTR to ablate candidate progenitor populations, assessing whether regeneration is subsequently impaired. The Raible lab used two complementary ablation approaches to pinpoint the cells responsible for regeneration [
40
]. Neomycin, an aminoglycoside antibiotic, reliably ablates mature hair cells [
39
], which normally regenerate fully. In parallel, the NTR/MTZ system was used to selectively ablate dorsoventral (DV) support cells, which were suspected to act as progenitors. When both the mature hair cells and DV support cells were eliminated, regeneration was dramatically impaired [
40
]. This dual-ablation strategy demonstrates another way the NTR/prodrug system can be used to identify the cell populations that contribute to regeneration.
NTR/Prodrug – Dependent Ablation in Modeling Human Disease
The NTR/prodrug ablation system also lends itself well to modeling human diseases that are characterized by the specific and progressive loss of distinct cell populations. The core strengths of this chemogenetic approach include cell-type specificity, and temporal control which allows it to be a toolkit for recapitulating pathological events
in vivo
. This allows for the real-time dissection of disease initiation, progression, and complex cellular responses to injury. A diverse array of human pathologies has been modeled using NTR/MTZ in zebrafish, including chronic hyperglycemia (a symptom of diabetes) [
120
], acute liver damage [
116
], and cardiac injury [
106
,
121
,
122
]. Here, we analyze selected models in greater detail, focusing on kidney disease, retinal degeneration, demyelinating disorders, and neurodegeneration. (
Figure 1
– orange boxes).
Kidney Glomerular Disease
Glomerular diseases stem from a common problem: progressive podocyte loss or dysfunction. These cells are crucial for maintaining the kidney’s filtration barrier and their damage leads to proteinuria, the leakage of abnormal amounts of protein into the urine [
123
]. To model this pathology, a podocyte-specific NTR mouse line was generated, in which NTR expression under the
podocin
promoter enables inducible podocyte injury [
92
]. Administration of CB1954 triggered acute podocyte damage with proteinuria and leads to progressive focal segmental glomerulosclerosis, providing a mammalian proof-of-principle for NTR/CB1954-mediated podocyte ablation, though this approach was never widely adopted. Subsequent mouse studies of podocyte ablation instead used DTR-based approaches [
124
–
126
], even though DT alone can induce transient podocyte injury and proteinuria in wild-type mice [
62
].
Other investigators employed the NTR/MTZ to induce podocyte-specific loss in zebrafish. Researchers used the
nphs2
promoter to drive NTR in transgenic zebrafish (nphs2:NTR-GFP), enabling precise, inducible podocyte ablation.
[
127
,
128
] Administering MTZ triggered rapid podocyte apoptosis, resulting in classic features of human glomerular injury, including disruption of the filtration barrier, proteinuria, and edema. By mirroring these essential aspects, this model provides a direct and relevant system for studying the progression of human podocytopathies. Furthermore, the zebrafish pronephros allows live imaging of podocyte injury and subsequent regeneration [
127
]. After MTZ withdrawal, podocyte repopulation occurs through residual cells and local progenitors [
128
]. This makes the model an ideal platform for studying podocyte repair and uncovering pathways with therapeutic relevance.
Retinal Degeneration
Given the high conservation of eye structure between zebrafish and humans, the NTR/MTZ system provides an excellent platform for modeling inherited retinal degenerations such as retinitis pigmentosa and cone dystrophies, conditions in which progressive photoreceptor loss leads to vision decline [
129
]. By driving NTR expression under photoreceptor-specific promoters, distinct photoreceptor subtypes can be selectively ablated. For example, expression under the rhodopsin (rho) promoter enables targeted elimination of rod photoreceptors, providing a robust zebrafish model of retinitis pigmentosa [
10
]. Similarly, the use of cone opsin promoters such as
opnlswl
permits ablation of defined cone populations to study cone dystrophies [
130
]. This targeted ablation triggers apoptotic photoreceptor loss while sparing neighboring retinal cells [
131
]. A major advantage of the zebrafish system is its capacity for spontaneous retinal regeneration, driven by the dedifferentiation and proliferation of Müller glia that give rise to new photoreceptors [
132
,
133
]. The NTR/MTZ paradigm allows for precise initiation and synchronization of this regenerative process, enabling realtime dissection of the cellular and molecular programs underlying photoreceptor replacement and the contributions of innate immune signaling to retinal repair [
83
].
Demyelinating Disorders
Conventional autoimmune models of Multiple Sclerosis (MS), including Experimental Autoimmune Encephalomyelitis (EAE), often display substantial variability in the timing and severity of disease onset, alongside a highly complex and multifactorial immunopathology [
134
]. This inherent heterogeneity makes it difficult to disentangle the individual cellular and molecular events that specifically contribute to successful remyelination, thereby limiting the ability to clearly define the mechanisms required for effective tissue repair. To overcome these hurdles, NTR-MTZ based models have been utilized to ablate oligodendrocytes and their progenitors. Tg(
mbp:gal4-vp16
); Tg(
UAS-E1B:NTR-mCherry
) fish express NTR specifically in mature oligodendrocytes that myelinate CNS axons. Exposure to MTZ in these fish caused rapid and synchronized demyelination within 48 hours, characterized by the retraction of myelin sheaths and oligodendrocyte cell death [
135
]. Furthermore, subsequent regeneration resulted in myelin sheaths that restored normal length and thickness correlated to axon caliber [
136
]. This mechanistic parallel is highly relevant, as the failure to restore proper myelin architecture is a central hallmark of progressive disability in human demyelinating diseases [
134
].
Dopaminergic Neurodegeneration
Traditional genetic models of Parkinson’s Disease (PD) often exhibit weak or late-onset phenotypes. Neurotoxin-based models frequently induce off-target neuronal loss, and the compounds themselves pose safety risks to researchers, restricting their use in scalable or high-content screening applications [
37
,
38
]. To overcome these hurdles, Kim
et al.
utilized a chemogenetic model in zebrafish to abate dopaminergic (DA) neurons [
109
] NTR expression was driven from the
tyrosine hydroxylase (th)
promo ter (the
th1
gene encodes an enzyme required for dopamine synthesis).
Th:NTR
fish express NTR1.0 in the DA neurons of the ventral forebrain, the zebrafish homolog of the mammalian substantia nigra.
Exposure to MTZ in
th:NTR
fish caused pronounced mitochondrial damage within DA neurons, including mtDNA damage, impaired mitochondrial function, reduced organelle motility, and altered morphology, ultimately resulting in neuron loss [
109
]. The finding that NTR/MTZ ablation kills dopaminergic neurons through mitochondrial dysfunction is particularly significant, as mitochondrial impairment is a central pathological hallmark of human PD [
137
]. By recapitulating this key feature, the
th:NTR
model moves beyond a simple cell-elimination system to one with strong disease relevance, providing a robust, scalable, and experimentally tractable platform ideally suited for screening small molecules that protect DA neurons or modulate PD-associated pathways. Furthermore, this work remains one of the few detailed mechanistic investigations of NTR/MTZ-mediated cytotoxicity in a defined zebrafish neuronal population, indicating that mitochondrial injury, rather than early nuclear DNA damage, plays a major role in driving NTR/MTZ-induced dopaminergic neuron death.
NTR/Prodrug – Based Screening
The NTR/MTZ ablation system provides a reproducible and scalable platform for functional screening in zebrafish, combining cell-type specificity, quantitative imaging, and compatibility with both chemical and genetic perturbations (
Figure 2
).
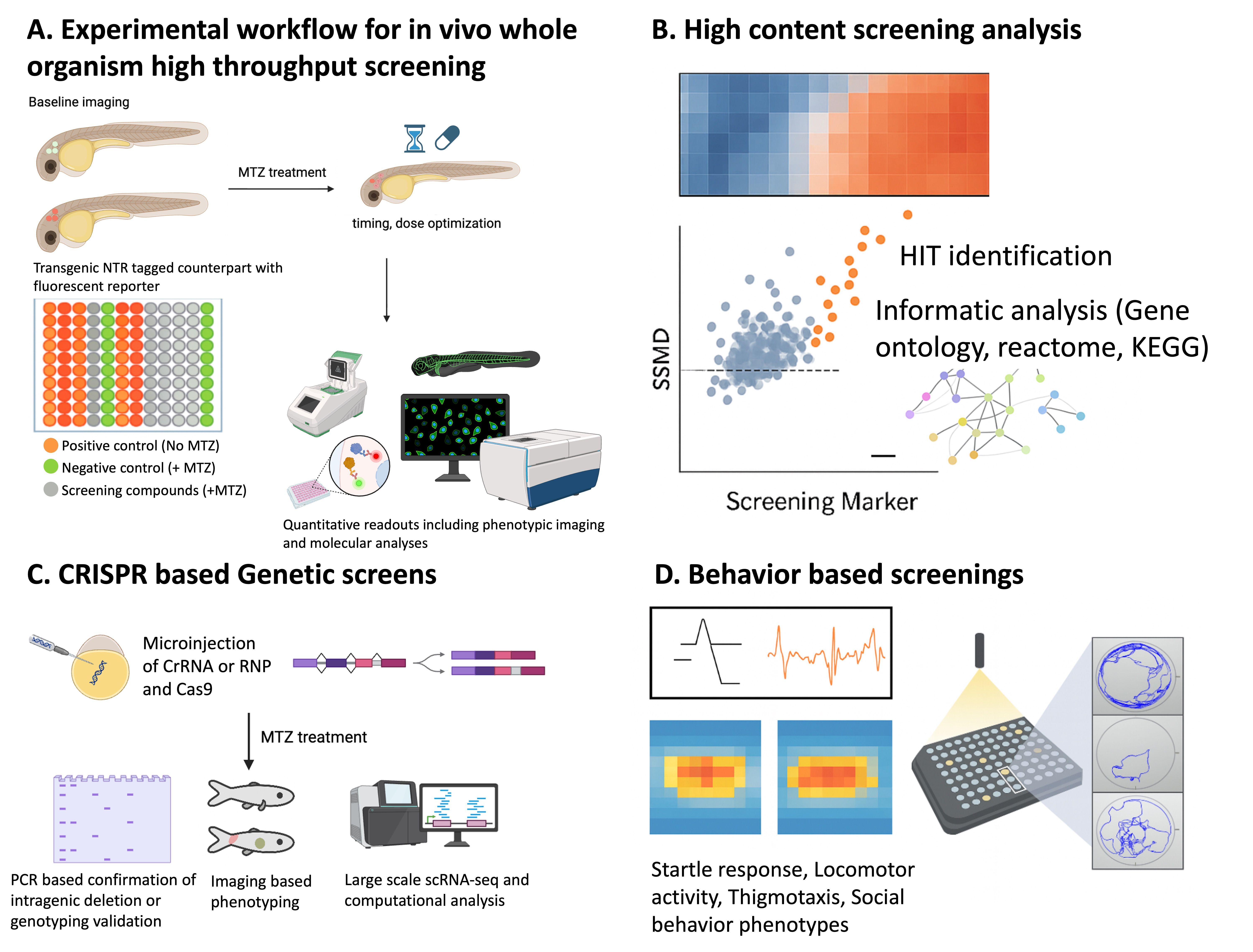
NTR/MTZ-based screening platforms in zebrafish.
Overview of the integrated chemogenetic screening workflow using nitroreductase (NTR)–mediated ablation. (A) Experimental design showing transgenic zebrafish expressing NTR in target tissues, baseline imaging, and subsequent metronidazole (MTZ) treatment to induce cell-type–specific ablation. The use of parallel transgenic controls and multiwell plate layout enables quantitative assessment of tissue loss and recovery. (B) High-content chemical screening pipeline integrating automated imaging, hit identification, and pathway-level analysis using standardized statistical metrics. (C) Genetic screening framework coupling sgRNA-based mutagenesis with imaging-based phenotype scoring to uncover modifiers of cell loss or regeneration. (D) Behavioral assays to quantify functional recovery or pharmacological response.
Small-Molecule Screening
The NTR/MTZ ablation system has been adapted for high-content chemical screening in zebrafish, allowing quantitative evaluation of compound effects on cell death, protection, and regeneration across diverse tissues. In the context of retinal degeneration [
138
], Mumm and colleagues demonstrated the high-throughput capabilities of the system by screening 2,934 compounds using the
Tg(rho:YFP-NTR)
model of retinitis pigmentosa. By driving NTR specifically in rod photoreceptors, the lab induced targeted cell death and screened for small molecules that could preserve YFP-positive cells despite MTZ exposure. This large-scale effort identified 11 validated neuroprotectants (distinct from simple antioxidants) that were subsequently shown to have conserved efficacy in mouse retinal explant assays [
138
]. This cross-species validation confirms that the zebrafish NTR system effectively filters for compounds with relevant translational potential for human blindness.
Kim
et al
. (2022) utilized the
Tg(th:NTR)
model to perform a 1,403-compound screen for Parkinson’s disease. By integrating automated imaging with rigorous statistical metrics including the Brain Health Score (BHS) and the Strictly Standardized Mean Difference (SSMD), the researchers identified 57 compounds that preserved dopaminergic neurons (
Figure 2A-B
). Importantly, the study advanced beyond simple measurements of cell survival and provided mechanistic validation that these compounds protected neurons by restoring mitochondrial function, which is a central hallmark of PD pathology. The predictive validity of these hits was further confirmed through cross-assay validation in a separate Gaucher disease behavior model, demonstrating the system’s capacity to identify robust therapeutics for complex neurodegenerative conditions (
Figure 2D
) [
109
].
Another promising application of the NTR system lies in the identification of therapeutic agents that actively promote tissue regeneration. Lee
et al.
(2025) leveraged an optimized QF-based binary expression system (
mbpa:qf2;quas:epNTR-P2A-mCherry
) to perform a remyelination phenotypic screen for regenerative compounds. This transgenic line achieved greater than 85% oligodendrocyte loss following treatment with 2 mM MTZ for 18 hours, creating a highly reproducible regenerative baseline. Using this platform to screen a kinase-inhibitor library, the authors identified the TGF-β receptor I inhibitor AZ-12601011 as a potent driver of remyelination [
139
]. Mechanistic validation revealed that this compound promotes repair by modulating microglial and progenitor activation, thereby confirming the system’s predictive validity for discovering clinically relevant restorative therapeutics that actively drive the reconstruction of functional tissue.
Similar regenerative screens have been successfully implemented in other tissues, such as the pancreas. Andersson
et al.
(2012) utilized the
Tg(ins:CFP-NTR)
line, crossed with a
Tg(ins:Kaede)
reporter to induce complete β-cell ablation and then monitor the formation of new β cells. This model was used in a high-content screen of approximately 7,000 small molecules to find compounds that would enhance regeneration of the insulin producing β cells [
140
]. This screen identified adenosine receptor agonists, specifically NECA, as potent stimulators of endocrine regeneration. Detailed mechanistic characterization revealed that NECA signals via the A2aa receptor to specifically enhance the proliferation of regenerating β-cells rather than neogenesis, a therapeutic pathway that was subsequently validated to restore normoglycemia in a streptozotocin-induced diabetic mouse model [
140
].
Genetic and CRISPR-Based Screening
Chemical screens can identify potential therapeutic reagents, though their molecular targets often remain unknown. A complementary approach is to perform reverse-genetic screens that integrate NTR-mediated ablation with CRISPR mutagenesis to identify genes affecting regeneration [
141
]. This mutagenesis is achieved by injecting Cas9 ribonucleoprotein (RNP) complexes multiplexed with several guide RNAs per target gene directly into NTR-transgenic embryos (
Figure 2C
). This FO ‘crispant’ strategy generates high-efficiency somatic mutations in the first generation [
142
], allowing researchers to induce cell-specific ablation with MTZ and immediately quantify the effect of gene disruption on regeneration without the delay of establishing stable mutant lines.
To identify regulators of Retinal Pigment Epithelium (RPE) repair, Lu
et
al. (2023) conducted a focused F0 CRISPR screen targeting 27 candidate genes in rpe65a:nfsB-eGFP larvae [
143
]. By injecting ribonucleoprotein (RNP) complexes containing three highly mutagenic guide RNAs per gene, they achieved high-efficiency somatic mutagenesis in FO injected fish. The NTR/MTZ system induced the synchronized, widespread degeneration of the RPE, which subsequently triggered the secondary loss of photoreceptors. This screen identified numerous regulators of regeneration and revealed a novel mechanism that regulates the infiltration of phagocytic cells required for clearance of debris and complete regeneration [
143
].
To find regulators of retinal ganglion cell (RGC) regeneration, Emmerich
et al
. (2024) performed a large-scale CRISPR screen on 100 genes. Using the isl2b:Gal4; UAS:YFP-NTR2.0 line for RGC ablation, they identified 18 effector genes comprising key transcription factors and signaling pathway components [
144
]. The screen revealed that inhibition of Ascl1a accelerated the regeneration of new RGC neurons.
Finally, the integration of F0 mutagenesis with automated imaging establishes a scalable framework for future genetic screens. The ‘ZebraReg’ platform utilizes a dual-transgenic line (tbx5a:CreERT2; myh7l:loxP-tagBFP-STOP-loxP-mCherry-NTR) that restricts NTR expression specifically to the heart ventricle [
122
]. Treatment with MTZ ablated approximately 97% of cardiomyocytes, triggering a robust regenerative response that typically restores the tissue within three days. By combining this precise injury model with F0 CRISPR mutagenesis followed by immediate phenotyping, the study demonstrates a proof-of-concept workflow to understand the genetic mechanisms of cardiac repair.
Caveats and Improvements to the NTR/MTZ System
The NTR/MTZ system is widely used for diverse applications, but its performance has varied between labs. Key issues include batch-to-batch and preparation variability of MTZ, the need for high MTZ doses (≈10 mM) that can cause off-target toxicity (e.g., developing brain [
145
], larval/adult intestine [
120
]), and differential susceptibility of some cell types to ablation [
110
]. Because ablation rate depends on both NTR activity and MTZ dose, researchers have pursued three complementary strategies to improve reproducibility and experimental interpretation: 1) increase NTR expression, 2) engineer higher-activity NTR mutants, and 3) identify more efficacious prodrugs that achieve effective killing at lower, less toxic concentrations. These iterative improvements are aimed at mitigating previous limitations and expanding the range of feasible experimental paradigms and are discussed next:
1. Increase NTR expression
: Strong, well-characterized promoters/enhancers (e.g., the zebrafish insulin promoter) [
107
] can drive high NTR expression, especially when present in multiple copies via Tol2-mediated transgenesis [
146
]. However, many cell-specific regulatory elements are weak, and maintaining multiple insertions is challenging and prone to genetic drift and intergenerational variability. An alternative is to use a bipartite system such as Gal4/UAS, [
147
,
148
] which can produce robust, amplified NTR expression even from single genomic insertion. With this approach, a cell-specific promoter drives a Gal4 transactivator that binds UAS sites to strongly activates
NTR
transcription (
Figure 3
). For example, elements from the 14xUAS constructs of Köster and Fraser [
148
] were used to generate the transgenic line Tg(UAS-ElB:NTR-mCherry)
c264
. [
110
,
149
] These fish were distributed by the Zebrafish International Resource Center (ZIRC) and have been widely used by the zebrafish community: 10 of the 32 most-cited papers on zebrafish nitroreductase ablation use this line (
Supplemental Table 1
). A caveat is that Gal4/UAS DNA elements can be prone to epigenetic silencing, producing mosaic expression; the repetitive UAS contains multiple CpG sites susceptible to DNA methylation. [
150
,
151
] Silencing can be mitigated by using a less repetitive UAS (e.g., 4x) [
152
] or by using the QF/QUAS bipartite system (derived from Neurospora), [
153
] which has been reported to show reduced silencing [
154
] and has recently been adapted for NTR-based ablation (
Figure 3
). [
139
,
155
]
2. Higher-activity NTR mutants
: Substantial effort has gone into engineering more active NTRs; first driven by their promise as cancer ‘suicide-gene’ therapies [
156
] and later to improve NTR-based ablation in basic research. [
157
] Two research groups independently engineered the same three substitutions into the wild-type
E. coli
enzyme (now termed NTR1.0), creating more efficient versions they named epNTR and NTR1.1. [
156
–
158
]. Cross-species screening identified a highly active nitroreductase (NTR) from
Vibrio vulnificus
. Using this enzyme as a scaffold, rational engineering yielded the second-generation variant NTR2.0, which exhibits a greater than 100-fold enhancement in activity over the original NTR1.0. [
78
]
The use of first-generation nitroreductase (NTR1) for chronic cell ablation was problematic, as the required 10 mM metronidazole (MTZ) dose induces intestinal pathology and approaches the LD50 in zebrafish. [
120
] However, the more active NTR2.0 variant enables effective ablation with far lower, better-tolerated MTZ concentrations. To demonstrate this, Tucker
et al
. developed a zebrafish model expressing NTR2.0 specifically in pancreatic β cells. [
152
] They found that efficient larval β cell ablation required only 100 μM MTZ, a regimen that could be maintained for 10 days without ill effects. In stark contrast, the NTR1 system required a toxic 10 mM MTZ dose, which is lethal to larvae (independent of NTR) within three days. In adult fish, a regimen of 5 mM MTZ for two days followed by two weeks at 1 mM was completely tolerated by wild-type fish with no ill effects but induced sustained hyperglycemia and weight loss in NTR2.0-expressing fish. This established a powerful model for studying chronic diabetic consequences, such as retinopathy, nephropathy, and impaired wound healing.
This well-tolerated ablation paradigm now makes it possible to model a range of other chronic conditions, including neurodegenerative, renal, and muscular disorders. This capability, in turn, facilitates the study of long-term disease progression and the evaluation of new therapeutic interventions.
3. More efficacious prodrugs
: Metronidazole (MTZ) efficacy can vary across suppliers and batches. To ensure consistency, it is recommended to prepare fresh MTZ solutions for experiments. [
110
,
120
] To overcome MTZ’s limitations, alternative prodrugs like nifurpirinol (NFP) have been tested. NFP is a more potent nitrofuran-based prodrug. [
110
,
120
] However, its structural class is distinct from the nitroimidazole-based prodrugs for which NTR2.0 was specifically engineered. [
78
] As a nitroimidazole prodrug, Ronidazole (RNZ) likely retains compatibility with newer NTR systems while offering significant practical advantages over MTZ, primarily its better potency. [
80
,
145
,
159
] For instance, in
Tg(fabpl 0:mCherry-NTRl
) fish, 2 mM RNZ achieved hepatocyte ablation comparable to 10 mM MTZ, a five-fold increase in potency. [
159
] This pattern of higher efficacy was replicated in a macrophage model, where a five-fold lower RNZ dose was as effective as MTZ. [
145
] Lai
et al
. also reported no bystander effects and demonstrated RNZ efficacy with the NTR1.1 variant. Recently, it has also been shown that RNZ functions with NTR2.0 to cause cell-specific ablation, [
160
] although a direct comparison of RNZ versus MTZ with NTR2.0 has not been reported. Whether RNZ shows batch-to-batch variability similar to MTZ has not yet been reported. Given the potential for variability, it would be prudent for researchers to titrate each new batch of RNZ or, alternatively, adopt a dosing strategy that exceeds the minimum effective concentration to ensure consistent ablation results. Nonetheless, it is anticipated that the NTR2.0/RNZ combination will further lower the required prodrug concentrations and minimize off-target activity.
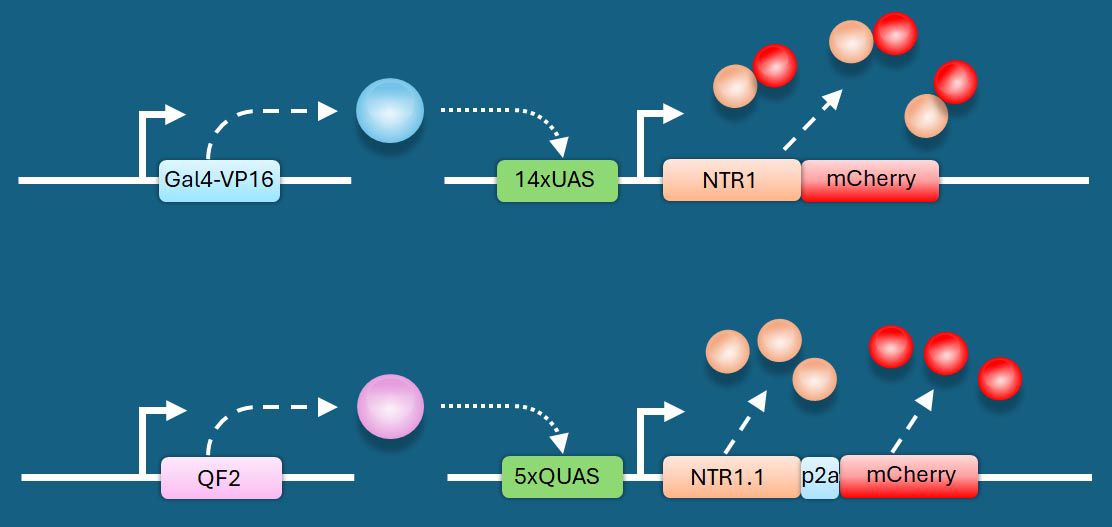
Schematic of bipartite systems to drive robust levels of NTR.
On left, driver lines express (dashed arrows) transactivators (A) Gal4 (blue sphere), or (B) QF2 (purple sphere) under the control of a cis-regulatory element (CRE). These transactivators bind their respective upstream activating sequences (either UAS or QUAS, grey boxes) to achieve controlled and amplified NTR expression (tan spheres) in target cells. NTR expression can be monitored by co-production of mCherry (red spheres) either as a fusion protein or as separate proteins due to P2A dependent ribosome ‘skipping’
162
. (A) redrawn from Pisharath & Parsons where the CRE was from ptf1a
110
, and (B) redrawn from Lee et al. where the CRE came from mbpa
139
.
Given this evolving landscape of prodrugs and enzymes, what are the critical factors a researcher must weigh when designing an NTR ablation experiment?
Experimental Design: Practical and Technical Considerations
A successful ablation experiment using the NTR/prodrug system requires careful consideration of three key components: (1) transgenic strategy, (2) optimal NTR activity, and (3) appropriate controls. The optimal design depends on the biological question.
◦ For regeneration studies, aim for complete ablation to clearly assess neogenesis.
◦ For functional studies, partial ablation may be sufficient to reveal a phenotype.
1.
Transgenic strategy:
Regulatory elements
: Select regulatory elements that ensure precise tissue or cell-type specificity.
Discrete regulation
: When a single promoter is insufficient to achieve the desired tissue specificity, use intersectional approaches (e.g., Cre/lox) to restrict expression. [
161
]
Fluorescent marker
: Include an independent fluorescent marker (fusion or 2A reporter [
162
]) to identify transgenic animals and confirm appropriate expression.
Positional effects
: NTR transgenes, like any transgene, can exhibit positional effects (leakiness, mosaicism). To ensure reliable lines:
Screen multiple founders
: Identify ≥5 FO founders and ideally establish 5 independent F1 lines.
Compare stable F1 lines
: Confirm that fluorescent-marker expression matches expected regulatory-element activity.
Prioritize F1 lines based on
:
– Mendelian transmission, indicating a single-site insertion.
– Consistent, non-mosaic expression of the fluorescent marker, indicating uniform NTR expression in all intended cells.
– Robust and reproducible expression, independent of whether the transgene is inherited maternally or paternally.
– Reliable and consistent ablation of target cell
2.
NTR activity:
General principle
: Strong NTR expression generally yields faster and more complete ablation. [
78
,
120
]
NTR variant
: NTR2.0 is currently the most active NTR variant used in zebrafish and is recommended for future studies.
Prodrug choice
: For most applications, RNZ is the recommended prodrug to start with, due to its higher efficacy and lower required dosing relative to MTZ, which can improve both ablation efficiency and experimental consistency.
Enhancing expression
: If the promoter driving NTR2.0 is weak, amplify expression using binary systems such as Gal4/UAS or QF/QUAS (
Figure 3
). [
139
,
149
,
155
]
3.
Controls:
Validating cell death
: To properly interpret ablation results, it is important to confirm that cell death occurred. Useful readouts include:
Apoptosis assays
: TUNEL, cleaved caspase-3 immunostaining. [
104
,
110
]
Loss of fluorescent reporters
: [
106
,
107
,
110
] If signal perdurance is a concern, use destabilized reporters to reduce fluorescence longevity. [
163
,
164
]
Cell-death kinetics
: As stressed cells may downregulate NTR and escape ablation, validating cell-death kinetics can be informative.
Use
endpoint analysis
(serial time-point fixation + apoptosis markers).
Or use
cell-death biosensors
, such as Hmgbl-GFP, which distinguishes necrosis (nuclear release) from apoptosis (nuclear retention). [
165
–
167
] Example: In larvae co-expressing
ins:mCherry-2A-NTR2.0
and
ins:hmgbl-eGFP
, [
120
,
168
] high-dose MTZ (1 mM) induced apoptosis within 4 hours with complete β-cell loss by 24 hours; a low dose (10 μM) produced slower dynamics (
Figure 4
).
Negative controls
NTR transgene, no prodrug
: Controls for effects of exogenous NTR expression.
No NTR transgene, prodrug
: Controls for nonspecific MTZ/RNZ effects, including antimicrobial activity. For microbiome-associated studies, consider alternative ablation methods.
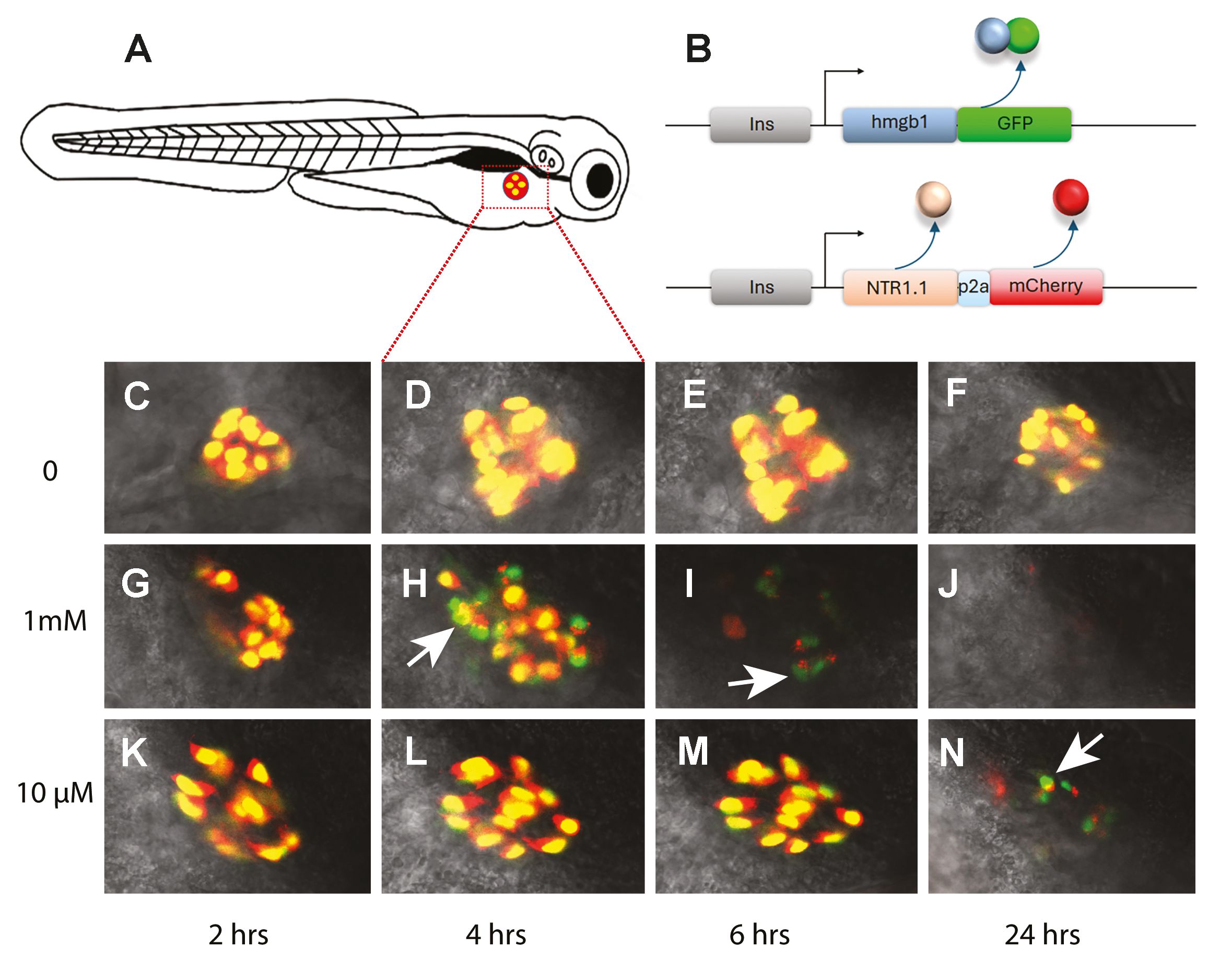
Live imaging of cell-death kinetics.
(A) Schematic of 6 dpf larvae showing position of the pancreatic islet imaged in C-N (red/yellow). (B) Diagram of the two transgenes (ins:Hmgb1-GFP, ins:mCherry-2a-NTR2.0) in the fish. The insulin promoter (grey box) drives expression of the following: (B-above) an Hmgb1-GFP fusion protein and (B-below) NTR2.0 and mCherry [presence of the P2A (blue box) makes separate proteins]. (C-N) Confocal images of the islet of in three larval fish over a time course from 6 dpf to 7 dpf (times along the X axis). (C-F) negative control – no MTZ (0). (G-J) fish treated with high MTZ dose (1mM). (K-N) fish treated with a low dose MTZ (10μM). (G-N) Dying β cells first lose red fluorescence, revealing green nuclei (arrow heads). A higher dose shows appearance of green nuclei (H) earlier than the lower dose (N). (J) 24 hrs in 1mM and no debris remains.
Additional files
References
1.
Advances in understanding tissue regenerative capacity and mechanisms in animals
Nat Rev Genet
11
:710–722
PubMed
Google Scholar
2.
The cellular basis for animal regeneration
Dev Cell
21
:172–185
PubMed
Google Scholar
3.
Regenerative tissue remodeling in planarians - The mysteries of morphallaxis
Semin Cell Dev Biol
87
:13–21
PubMed
Google Scholar
4.
Wound repair and regeneration
Nature
453
:314–321
PubMed
Google Scholar
5.
Dedifferentiation, transdifferentiation and reprogramming: three routes to regeneration
Nat Rev Mol Cell Biol
12
:79–89
PubMed
Google Scholar
6.
Zebrafish disease models in drug discovery: from preclinical modelling to clinical trials
Nat Rev Drug Discov
20
:611–628
PubMed
Google Scholar
7.
Heart regeneration in zebrafish
Science
298
:2188–2190
PubMed
Google Scholar
8.
Hooked on heart regeneration: the zebrafish guide to recovery
Cardiovasc Res
118
:1667–1679
PubMed
Google Scholar
9.
Visual Function is Gradually Restored During Retina Regeneration in Adult Zebrafish
Front Cell Dev Biol
9
:831322
PubMed
Google Scholar
10.
A novel model of retinal ablation demonstrates that the extent of rod cell death regulates the origin of the regenerated zebrafish rod photoreceptors
J Comp Neurol
518
:800–814
PubMed
Google Scholar
11.
Axonal regrowth after spinal cord transection in adult zebrafish
J Comp Neurol
377
:577–595
https://doi.org/10.1002/(sici)1096-9861(19970127)377:4<577::aid-cne8>3.0.co;2-#
PubMed
Google Scholar
12.
Progenitor-derived glia are required for spinal cord regeneration in zebrafish
Development
150
PubMed
Google Scholar
13.
The regenerative capacity of the zebrafish caudal fin is not affected by repeated amputations
PLoS One
6
:e22820
PubMed
Google Scholar
14.
Zebrafish fin regeneration involves generic and regeneration-specific osteoblast injury responses
eLife
11
PubMed
Google Scholar
15.
Model systems for regeneration: zebrafish
Development
146
PubMed
Google Scholar
16.
The art of fin regeneration in zebrafish
Regeneration
2
:72–83
PubMed
Google Scholar
17.
Regeneration of the adult zebrafish brain from neurogenic radial glia-type progenitors
Development
138
:4831–4841
PubMed
Google Scholar
18.
In Toto Imaging of Dynamic Osteoblast Behaviors in Regenerating Skeletal Bone
Curr Biol
28
:3937–3947
PubMed
Google Scholar
19.
Regenerating zebrafish scales express a subset of evolutionary conserved genes involved in human skeletal disease
BMC Biol
20
:21
PubMed
Google Scholar
20.
Endogenous tenocyte activation underlies the regenerative capacity of the adult zebrafish tendon
NPJ Regen Med
8
:52
PubMed
Google Scholar
21.
Ligament injury in adult zebrafish triggers ECM remodeling and cell dedifferentiation for scar-free regeneration
bioRxiv
PubMed
Google Scholar
22.
Motor neuron regeneration in adult zebrafish
J Neurosci
28
:8510–8516
PubMed
Google Scholar
23.
Fgf-dependent glial cell bridges facilitate spinal cord regeneration in zebrafish
J Neurosci
32
:7477–7492
PubMed
Google Scholar
24.
Cryoinjury as a myocardial infarction model for the study of cardiac regeneration in the zebrafish
Nat Protoc
7
:782–788
PubMed
Google Scholar
25.
Multiple cryoinjuries modulate the efficiency of zebrafish heart regeneration
Sci Rep
10
:11551
PubMed
Google Scholar
26.
Light-induced rod and cone cell death and regeneration in the adult albino zebrafish (Danio rerio) retina
J Neurobiol
44
:289–307
https://doi.org/10.1002/1097-4695(20000905)44:3<289::aid-neu1>3.0.co;2-h
PubMed
Google Scholar
27.
Cell proliferation follows acoustically-induced hair cell bundle loss in the zebrafish saccule
Hear Res
253
:67–76
PubMed
Google Scholar
28.
The stat3/socs3a pathway is a key regulator of hair cell regeneration in zebrafish. [corrected]
J Neurosci
32
:10662–10673
PubMed
Google Scholar
29.
Larval Zebrafish Lateral Line as a Model for Acoustic Trauma
eNeuro
5
PubMed
Google Scholar
30.
Laser ablations reveal functional relationships of segmental hindbrain neurons in zebrafish
Neuron
23
:325–335
https://doi.org/10.1016/s0896-6273(00)80783-7
PubMed
Google Scholar
31.
Ablation of a Neuronal Population Using a Two-photon Laser and Its Assessment Using Calcium Imaging and Behavioral Recording in Zebrafish Larvae
J Vis Exp
PubMed
32.
Visuomotor behaviors in larval zebrafish after GFP-guided laser ablation of the optic tectum
J Neurosci
23
:3726–3734
PubMed
Google Scholar
33.
Laser-targeted ablation of the zebrafish embryonic ventricle: a novel model of cardiac injury and repair
Int J Cardiol
168
:3913–3919
PubMed
Google Scholar
34.
Distinct inflammatory and wound healing responses to complex caudal fin injuries of larval zebrafish
eLife
8
PubMed
Google Scholar
35.
Damage-induced basal epithelial cell migration modulates the spatial organization of redox signaling and sensory neuron regeneration
eLife
13
PubMed
Google Scholar
36.
Zebrafish macrophages convert physical wound signals into rapid vascular permeabilization
Nat Commun
17
:1807
PubMed
Google Scholar
37.
Animal models of Parkinson’s disease: bridging the gap between disease hallmarks and research questions
Transl Neurodegener
12
:36
PubMed
Google Scholar
38.
MPTP mouse models of Parkinson’s disease: an update
J Parkinsons Dis
1
:19–33
PubMed
Google Scholar
39.
Bax, Bcl2, and p53 differentially regulate neomycin-and gentamicin-induced hair cell death in the zebrafish lateral line
J Assoc Res Otolaryngol
14
:645–659
PubMed
Google Scholar
40.
Distinct progenitor populations mediate regeneration in the zebrafish lateral line
eLife
8
PubMed
Google Scholar
41.
Aminoglycoside-induced hair cell death of inner ear organs causes functional deficits in adult zebrafish (Danio rerio)
PLoS One
8
:e58755
PubMed
Google Scholar
42.
PI3K and Inhibitor of Apoptosis Proteins Modulate Gentamicin-Induced Hair Cell Death in the Zebrafish Lateral Line
Front Cell Neurosci
11
:326
PubMed
Google Scholar
43.
Regeneration of the pancreas in adult zebrafish
Diabetes
58
:1844–1851
PubMed
Google Scholar
44.
PGE2-regulated wnt signaling and N-acetylcysteine are synergistically hepatoprotective in zebrafish acetaminophen injury
Proc Natl Acad Sci USA
107
:17315–17320
PubMed
Google Scholar
45.
A rapid protocol for inducing acute pancreatitis in zebrafish models
Comp Biochem Physiol C Toxicol Pharmacol
283
:109958
PubMed
Google Scholar
46.
Cerulein pancreatitis: oxidative stress, inflammation, and apoptosis
Gut Liver
2
:74–80
PubMed
Google Scholar
47.
Direct targeting of mitochondria by cisplatin leads to cytotoxicity in zebrafish lateral-line hair cells
iScience
27
:110975
PubMed
Google Scholar
48.
Mechanical overstimulation causes acute injury and synapse loss followed by fast recovery in lateral-line neuromasts of larval zebrafish
eLife
10
PubMed
Google Scholar
49.
Regeneration of inner retinal neurons after intravitreal injection of ouabain in zebrafish
J Neurosci
27
:1712–1724
PubMed
Google Scholar
50.
Ganglion cell regeneration following whole-retina destruction in zebrafish
Dev Neurobiol
68
:166–181
PubMed
Google Scholar
51.
Small molecule-induced ablation and subsequent regeneration of larval zebrafish melanocytes
Development
133
:3563–3573
PubMed
Google Scholar
52.
Non-neuromodulatory Optogenetic Tools in Zebrafish
Front Cell Dev Biol
8
:418
PubMed
Google Scholar
53.
Seeing the Light: The Use of Zebrafish for Optogenetic Studies of the Heart
Front Physiol
12
:748570
PubMed
Google Scholar
54.
Precise spatio-temporal control of rapid optogenetic cell ablation with mem-KillerRed in Zebrafish
Sci Rep
7
:5096
PubMed
Google Scholar
55.
A genetically encoded photosensitizer
Nat Biotechnol
24
:95–99
PubMed
Google Scholar
56.
Optogenetic in vivo cell manipulation in KillerRed-expressing zebrafish transgenics
BMC Dev Biol
10
:110
PubMed
Google Scholar
57.
Real-time visualization of oxidative stress-mediated neurodegeneration of individual spinal motor neurons in vivo
Redox Biol
19
:226–234
PubMed
Google Scholar
58.
In vivo optogenetics for light-induced oxidative stress in transgenic zebrafish expressing the KillerRed photosensitizer protein
Methods Mol Biol
1148
:229–238
PubMed
Google Scholar
59.
Understanding the mode of action of diphtheria toxin: a perspective on progress during the 20th century
Toxicon
39
:1793–1803
https://doi.org/10.1016/s0041-0101(01)00165-9
PubMed
Google Scholar
60.
Expression cloning of a diphtheria toxin receptor: identity with a heparin-binding EGF-like growth factor precursor
Cell
69
:1051–1061
https://doi.org/10.1016/0092-8674(92)90623-k
PubMed
Google Scholar
61.
Diphtheria toxin receptor-mediated conditional and targeted cell ablation in transgenic mice
Nat Biotechnol
19
:746–750
PubMed
Google Scholar
62.
Impairment of podocyte function by diphtheria toxin--a new reversible proteinuria model in mice
Lab Invest
92
:1674–1685
PubMed
Google Scholar
63.
Vestibular and Auditory Hair Cell Regeneration Following Targeted Ablation of Hair Cells With Diphtheria Toxin in Zebrafish
Front Cell Neurosci
15
:721950
PubMed
Google Scholar
64.
ptf1a(+), ela31(-) cells are developmentally maintained progenitors for exocrine regeneration following extreme loss of acinar cells in zebrafish larvae
Dis Model Mech
10
:307321
PubMed
Google Scholar
65.
Suppression of lens growth by alphaA-crystallin promoter-driven expression of diphtheria toxin results in disruption of retinal cell organization in zebrafish
Dev Biol
255
:113–127
https://doi.org/10.1016/s0012-1606(02)00079-9
PubMed
Google Scholar
66.
DTA-mediated targeted ablation revealed differential interdependence of endocrine cell lineages in early development of zebrafish pancreas
Differentiation
78
:241–252
PubMed
Google Scholar
67.
A Genetic Cardiomyocyte Ablation Model for the Study of Heart Regeneration in Zebrafish
Methods Mol Biol
2158
:71–80
PubMed
Google Scholar
68.
Systematic optimization and prediction of cre recombinase for precise genome editing in mice
Genome Biol
26
:85
PubMed
Google Scholar
69.
Caspases: intracellular signaling by proteolysis
Cell
91
:443–446
https://doi.org/10.1016/s0092-8674(00)80430-4
PubMed
Google Scholar
70.
Mechanisms of caspase activation
Curr Opin Cell Biol
15
:725–731
PubMed
Google Scholar
71.
The apoptosome: signalling platform of cell death
Nat Rev Mol Cell Biol
8
:405413
PubMed
Google Scholar
72.
A rapid, reversible, and tunable method to regulate protein function in living cells using synthetic small molecules
Cell
126
:9951004
PubMed
Google Scholar
73.
Lifelong regeneration of cerebellar Purkinje cells after induced cell ablation in zebrafish
eLife
12
PubMed
Google Scholar
74.
Caspase-mediated apoptosis induction in zebrafish cerebellar Purkinje neurons
Development
143
:4279–4287
PubMed
Google Scholar
75.
Novel caspase-suicide proteins for tamoxifen-inducible apoptosis
Genesis
46
:530–536
PubMed
Google Scholar
76.
Appropriate subcellular localisation of prodrug-activating enzymes has important consequences for suicide gene therapy
Int J Cancer
93
:123–130
PubMed
Google Scholar
77.
Analysis of beta cell proliferation dynamics in zebrafish
Dev Biol
332
:299–308
PubMed
Google Scholar
78.
NTR 2.0: a rationally engineered prodrug-converting enzyme with substantially enhanced efficacy for targeted cell ablation
Nat Methods
19
:205–215
PubMed
Google Scholar
79.
Selective Cell Ablation Using an Improved Prodrug-Converting Nitroreductase
Methods Mol Biol
2707
:223–234
PubMed
Google Scholar
80.
Spatiotemporal control of cell ablation using Ronidazole with Nitroreductase in Drosophila
Dev Biol
520
:31–40
PubMed
Google Scholar
81.
Ectopic head regeneration after nervous system ablation in a sea anemone
Curr Biol
35
:5955–5964
PubMed
Google Scholar
82.
Live imaging of targeted cell ablation in Xenopus: a new model to study demyelination and repair
J Neurosci
32
:12885–12895
PubMed
Google Scholar
83.
Muller glial cell reactivation in Xenopus models of retinal degeneration
Glia
65
:1333–1349
PubMed
Google Scholar
84.
The Xenopus tadpole: An in vivo model to screen drugs favoring remyelination
Mult Scler
24
:1421–1432
PubMed
Google Scholar
85.
Rod-Specific Ablation Using the Nitroreductase/Metronidazole System to Investigate Regeneration in Xenopus
Cold Spring Harb Protoc
2018
PubMed
Google Scholar
86.
Conditional ablation of osteoblasts in medaka
Dev Biol
364
:128–137
PubMed
Google Scholar
87.
Ablation of central nervous system progenitor cells in transgenic rats using bacterial nitroreductase system
J Neurosci Res
85
:1183–1193
PubMed
Google Scholar
88.
Selective cell ablation in transgenic mice expression E. coli nitroreductase
Gene Ther
4
:101110
PubMed
Google Scholar
89.
The expression of bacterial nitroreductase in transgenic mice results in specific cell killing by the prodrug CB1954
Gene Ther
4
:93–100
PubMed
Google Scholar
90.
Inducible ablation of adipocytes in adult transgenic mice expressing the E. coli nitroreductase gene
J Endocrinol
175
:487–498
PubMed
Google Scholar
91.
Conditional ablation of neurones in transgenic mice
J Neurobiol
47
:183–193
PubMed
Google Scholar
92.
Transgenic mice expressing nitroreductase gene under the control of the podocin promoter: a new murine model of inductible glomerular injury
Virchows Arch
456
:325–337
PubMed
Google Scholar
93.
Type I nitroreductases of Escherichia coli
Can J Microbiol
27
:81–86
PubMed
Google Scholar
94.
Oxygen-insensitive nitroreductases: analysis of the roles of nfsA and nfsB in development of resistance to 5-nitrofuran derivatives in Escherichia coli
J Bacteriol
180
:5529–5539
PubMed
Google Scholar
95.
Genetics of nitrofurazone resistance in Escherichia coli
J Bacteriol
133
:10–16
PubMed
Google Scholar
96.
Prodrugs: a challenge for the drug development
Pharmacol Rep
65
:1–14
https://doi.org/10.1016/s1734-1140(13)70959-9
PubMed
Google Scholar
97.
Progress and problems with the use of suicide genes for targeted cancer therapy
Adv Drug Deliv Rev
99
:113–128
PubMed
Google Scholar
98.
The bioactivation of CB 1954 and its use as a prodrug in antibody-directed enzyme prodrug therapy (ADEPT)
Cancer Metastasis Rev
12
:195–212
PubMed
Google Scholar
99.
The nitroreductase enzyme in Walker cells that activates 5-(aziridin-1-yl)-2,4-dinitrobenzamide (CB 1954) to 5-(aziridin-l-yl)-4-hydroxylamino-2-nitrobenzamide is a form of NAD(P)H dehydrogenase (quinone) (EC )
Biochem Pharmacol
37
:4671–4677
https://doi.org/10.1016/0006-2952(88)90336-x
PubMed
Google Scholar
100.
Bioactivation of dinitrobenzamide mustards by an E. coli B nitroreductase
Biochem Pharmacol
50
:609–618
https://doi.org/10.1016/0006-2952(95)00187-5
PubMed
Google Scholar
101.
Virus-directed enzyme prodrug therapy: intratumoral administration of a replicationdeficient adenovirus encoding nitroreductase to patients with resectable liver cancer
J Clin Oncol
22
:1546–1552
PubMed
Google Scholar
102.
A phase I/II clinical trial in localized prostate cancer of an adenovirus expressing nitroreductase with CB1954 [correction of CB1984]
Mol Ther
17
:1292–1299
PubMed
Google Scholar
103.
The bystander effect of the nitroreductase/CB1954 enzyme/prodrug system is due to a cell-permeable metabolite
Hum Gene Ther
8
:709–717
PubMed
Google Scholar
104.
Nitroreductase-mediated cell/tissue ablation in zebrafish: a spatially and temporally controlled ablation method with applications in developmental and regeneration studies
Nat Protoc
3
:948–954
PubMed
Google Scholar
105.
Engineering the Escherichia coli Nitroreductase NfsA to Create a Flexible Enzyme-Prodrug Activation System
Front Pharmacol
12
:701456
PubMed
Google Scholar
106.
Conditional targeted cell ablation in zebrafish: a new tool for regeneration studies
Dev Dyn
236
:1025–1035
PubMed
Google Scholar
107.
Targeted ablation of beta cells in the embryonic zebrafish pancreas using E. coli nitroreductase
Mech Dev
124
:218–229
PubMed
Google Scholar
108.
Live imaging of leukocyte recruitment in a zebrafish model of chemical liver injury
Sci Rep
9
:28
PubMed
Google Scholar
109.
A zebrafish screen reveals Renin-angiotensin system inhibitors as neuroprotective via mitochondrial restoration in dopamine neurons
eLife
10
PubMed
Google Scholar
110.
Nitroreductase-mediated cell ablation in transgenic zebrafish embryos
Methods Mol Biol
546
:133–143
PubMed
Google Scholar
111.
Macrophages modulate adult zebrafish tail fin regeneration
Development
141
:2581–2591
PubMed
Google Scholar
112.
The Role of Macrophages During Zebrafish Injury and Tissue Regeneration Under Infectious and Non-Infectious Conditions
Front Immunol
12
:707824
PubMed
Google Scholar
113.
Centroacinar Cells Are Progenitors That Contribute to Endocrine Pancreas Regeneration
Diabetes
64
:3499–3509
PubMed
Google Scholar
114.
Centroacinar cells: At the center of pancreas regeneration
Dev Biol
413
:8–15
PubMed
Google Scholar
115.
Progenitor potential of nkx6.l-expressing cells throughout zebrafish life and during beta cell regeneration
BMC Biol
13
:70
PubMed
Google Scholar
116.
Extensive conversion of hepatic biliary epithelial cells to hepatocytes after near total loss of hepatocytes in zebrafish
Gastroenteroglogy
146
:776–788
PubMed
Google Scholar
117.
Regeneration of liver after extreme hepatocyte loss occurs mainly via biliary transdifferentiation in zebrafish
Gastroenterology
146
:789–800
PubMed
Google Scholar
118.
Decoding pancreatic endocrine cell differentiation and beta cell regeneration in zebrafish
Sci Adv
9
:eadf5142
PubMed
Google Scholar
119.
Cholangiocytes contribute to hepatocyte regeneration after partial liver injury during growth spurt in zebrafish
Nat Commun
16
:5260
PubMed
Google Scholar
120.
An inducible mode 1 of chronic hyperglycemia
Dis Model Mech
16
PubMed
Google Scholar
121.
Myocardium and BMP signaling are required for endocardial differentiation
Development
142
:2304–2315
PubMed
Google Scholar
122.
ZebraReg-a novel platform for discovering regulators of cardiac regeneration using zebrafish
Front Cell Dev Biol
12
:1384423
PubMed
Google Scholar
123.
Cell biology and pathology of podocytes
Annu Rev Physiol
74
:299–323
PubMed
Google Scholar
124.
Podocyte depletion causes glomerulosclerosis: diphtheria toxin-induced podocyte depletion in rats expressing human diphtheria toxin receptor transgene
J Am Soc Nephrol
16
:2941–2952
PubMed
Google Scholar
125.
Following specific podocyte injury captopril protects against progressive long term renal damage
F1000Res
4
:172
PubMed
Google Scholar
126.
Vascular Endothelial Growth Factor-A165b Restores Normal Glomerular Water Permeability in a Diphtheria-Toxin Mouse Model of Glomerular Injury
Nephron
139
:51–62
PubMed
Google Scholar
127.
Inducible podocyte injury and proteinuria in transgenic zebrafish
J Am Soc Nephrol
23
:1039–1047
PubMed
Google Scholar
128.
A zebrafish model of conditional targeted podocyte ablation and regeneration
Kidney Int
83
:1193–1200
PubMed
Google Scholar
129.
A review of the mechanisms of cone degeneration in retinitis pigmentosa
Acta Ophthalmol
94
:748–754
PubMed
Google Scholar
130.
Regeneration of cone photoreceptors when cell ablation is primarily restricted to a particular cone subtype
PLoS One
8
:e55410
PubMed
Google Scholar
131.
Immunomodulation-accelerated neuronal regeneration following selective rod photoreceptor cell ablation in the zebrafish retina
Proc Natl Acad Sci USA
114
:E3719-E3728
PubMed
Google Scholar
132.
Late-stage neuronal progenitors in the retina are radial Muller glia that function as retinal stem cells
J Neurosci
27
:7028–7040
PubMed
Google Scholar
133.
A role for alpha1 tubulin-expressing Muller glia in regeneration of the injured zebrafish retina
J Neurosci
26
:6303–6313
PubMed
Google Scholar
134.
Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS)
Br J Pharmacol
164
:1079–1106
PubMed
Google Scholar
135.
Generation of demyelination models by targeted ablation of oligodendrocytes in the zebrafish CNS
Mol Cells
36
:82–87
PubMed
Google Scholar
136.
Regeneration of myelin sheaths of normal length and thickness in the zebrafish CNS correlates with growth of axons in caliber
PLoS One
12
:e0178058
PubMed
Google Scholar
137.
Mitochondrial dysfunction in Parkinson’s disease - a key disease hallmark with therapeutic potential
Mol Neurodegener
18
:83
PubMed
Google Scholar
138.
Large-scale phenotypic drug screen identifies neuroprotectants in zebrafish and mouse models of retinitis pigmentosa
eLife
10
PubMed
Google Scholar
139.
Transforming growth factor-beta receptor I kinase plays a crucial role in oligodendrocyte regeneration after demyelination
BiomedPharmacother
187
:118094
PubMed
Google Scholar
140.
Adenosine signaling promotes regeneration of pancreatic beta cells in vivo
Cell Metab
15
:885–894
PubMed
Google Scholar
141.
Multiplexed CRISPR/Cas9 Targeting of Genes Implicated in Retinal Regeneration and Degeneration
Front Cell Dev Biol
6
:88
PubMed
Google Scholar
142.
A Rapid Method for Directed Gene Knockout for Screening in G0 Zebrafish
Dev Cell
46
:112–125
PubMed
Google Scholar
143.
A CRISPR-Cas9-mediated F0 screen to identify pro-regenerative genes in the zebrafish retinal pigment epithelium
Sci Rep
13
:3142
PubMed
Google Scholar
144.
A large-scale CRISPR screen reveals context-specific genetic regulation of retinal ganglion cell regeneration
Development
151
PubMed
Google Scholar
145.
Chemical screening reveals Ronidazole is a superior prodrug to Metronidazole for nitroreductase-induced cell ablation system in zebrafish larvae
J Genet Genomics
48
:1081–1090
PubMed
Google Scholar
146.
Rapid generation of single-insertion transgenics by Tol2 transposition in zebrafish
Dev Dyn
253
:1056–1065
PubMed
Google Scholar
147.
Targeted gene expression as a means of altering cell fates and generating dominant phenotypes
Development
118
:401–415
PubMed
Google Scholar
148.
Tracing transgene expression in living zebrafish embryos
Dev Biol
233
:329346
PubMed
Google Scholar
149.
Transactivation from Gal4-VP16 transgenic insertions for tissue-specific cell labeling and ablation in zebrafish
Dev Biol
304
:811–824
PubMed
Google Scholar
150.
Gal4/UAS transgenic tools a nd their application to zebrafish
Zebrafish
5
:97–110
PubMed
Google Scholar
151.
Transcriptional silencing and reactivation in transgenic zebrafish
Genetics
182
:747–755
PubMed
Google Scholar
152.
Transgenerational analysis of transcriptional silencing in zebrafish
Dev Biol
352
:191–201
PubMed
Google Scholar
153.
The Q system: a repressible binary system for transgene expression, lineage tracing, and mosaic analysis
Cell
141
:536–548
PubMed
Google Scholar
154.
Adoption of the Q transcriptional regulatory system for zebrafish transgenesis
Methods
66
:433–440
PubMed
Google Scholar
155.
The G-protein coupled receptor OXER1 is a tissue redox sensor essential for intestinal epithelial barrier integrity
bioRxiv
PubMed
Google Scholar
156.
Direct positive selection for improved nitroreductase variants using SOS triggering of bacteriophage lambda lytic cycle
Gene Ther
14
:690–698
PubMed
Google Scholar
157.
Enhanced cell-specific ablation in zebrafish using a triple mutant of Escherichia coli nitroreductase
Zebrafish
11
:85–97
PubMed
Google Scholar
158.
Direct activation of the Mauthner cell by electric field pulses drives ultrarapid escape responses
J Neurophysiol
112
:834–844
PubMed
Google Scholar
159.
Ronidazole Is a Superior Prodrug to Metronidazole for Nitroreductase-Mediated Hepatocytes Ablation in Zebrafish Larvae
Zebrafish
20
:95–102
PubMed
Google Scholar
160.
Chemogenetic ablation and regeneration of arterial valve in zebrafish
Biochem Biophys Res Commun
762
:151786
PubMed
Google Scholar
161.
Improved Tol2-mediated enhancer trap identifies weakly expressed genes during liver and beta cell development and regeneration in zebrafish
J Biol Chem
294
:932–940
PubMed
Google Scholar
162.
Viral 2A peptides allow expression of multiple proteins from a single ORF in transgenic zebrafish embryos
Genesis
45
:625–629
PubMed
Google Scholar
163.
Automated reporter quantification in vivo: high-throughput screening method for reporter-based assays in zebrafish
PLoS One
7
:e29916
PubMed
Google Scholar
164.
First quantitative high-throughput screen in zebrafish identifies novel pathways for increasing pancreatic beta-cell mass
eLife
4
PubMed
Google Scholar
165.
HMGB1: a signal of necrosis
Autoimmunity
40
:285–289
PubMed
Google Scholar
166.
Release of chromatin protein HMGB1 by necrotic cells triggers inflammation
Nature
418
:191–195
PubMed
Google Scholar
167.
Fluorescent biosensors for the detection of HMGB1 release
Methods Mol Biol
1004
:43–56
PubMed
Google Scholar
168.
Notch-responsive cells initiate the secondary transition in larval zebrafish pancreas
Mech Dev
126
:898–912
PubMed
Google Scholar
169.
Spinal motor neurons are regenerated after mechanical lesion and genetic ablation in larval zebrafish
Development
143
:1464–1474
PubMed
Google Scholar
170.
Gfap-positive radial glial cells are an essential progenitor population for later-born neurons and glia in the zebrafish spinal cord
Glia
64
:1170–1189
PubMed
Google Scholar
171.
Labelling and targeted ablation of specific bipolar cell types in the zebrafish retina
BMC Neurosci
10
:107
PubMed
Google Scholar
172.
Cellular dynamics of regeneration reveals role of two distinct Pax7 stem cell populations in larval zebrafish muscle repair
Dis Model Mech
9
:671–684
PubMed
Google Scholar
173.
Chemogenetic ablation of dopaminergic neurons leads to transient locomotor impairments in zebrafish larvae
J Neurochem
135
:249–260
PubMed
Google Scholar
174.
Establishment of a transgenic zebrafish line for superficial skin ablation and functional validation of apoptosis modulators in vivo
PLoS One
6
:e20654
PubMed
Google Scholar
175.
Regeneration of the zebrafish retinal pigment epithelium after widespread genetic ablation
PLoS Genet
15
:e1007939
PubMed
Google Scholar
176.
Sufficient numbers of early germ cells are essential for female sex development in zebrafish
PLoS One
10
:e0117824
PubMed
Google Scholar
177.
Inducible male infertility by targeted cell ablation in zebrafish testis
Mar Biotechnol
12
:466–478
PubMed
Google Scholar
178.
Nitroreductase-mediated gonadal dysgenesis for infertility control of genetically modified zebrafish
Mar Biotechnol
12
:569–578
PubMed
Google Scholar
179.
A transgenic zebrafish model of targeted oocyte ablation and de novo oogenesis
Dev Dyn
240
:1929–1937
PubMed
Google Scholar
180.
An In Vivo Zebrafish Model for Interrogating ROS-Mediated Pancreatic beta-Cell Injury, Response, and Prevention
Oxid Med Cell Longev
2018
:1324739
PubMed
Google Scholar
181.
Pineal photoreceptor cells are required for maintaining the circadian rhythms of behavioral visual sensitivity in zebrafish
PLoS One
7
:e40508
PubMed
Google Scholar
182.
Transparent adult zebrafish as a tool for in vivo transplantation analysis
Cell Stem Cell
2
:183–189
PubMed
Google Scholar
183.
Ablation of mpeg+ Macrophages Exacerbates mfrp-Related Hyperopia
Invest Ophthalmol Vis Sci
62
:13
PubMed
Google Scholar
184.
Tendon Cell Regeneration Is Mediated by Attachment Site-Resident Progenitors and BMP Signaling
Curr Biol
30
:3277–3292
PubMed
Google Scholar
Article and author information
Author information
Version history
Sent for peer review
:
Preprint posted
:
Reviewed Preprint version 1
:
Reviewed Preprint version 2
:
Cite all versions
You can cite all versions using the DOI
10.7554/eLife.110593
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2026,
Gha-Hyun J Kim & Michael J Parsons
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
views
315
downloads
17
citations
0
Views, downloads and citations are aggregated across all versions of this paper published by eLife.





