 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库Modulation of human dorsal root ganglion neuron excitability by Nav1.7 inhibition
Abstract
Nav1.7 voltage-gated sodium channels are strongly expressed in human primary painsensing neurons (nociceptors) and selective Nav1.7 inhibitors have been developed as possible therapeutic agents for treating pain, so far with disappointing clinical results. In contrast, a selective Nav1.8 channel inhibitor (suzetrigine) has had successful clinical trials. Because nociceptors express both Nav1.7 and Nav1.8 channels, it is of interest to compare effects of Nav1.7 and Nav1.8 inhibitors on the excitability of human nociceptors. To compare with previous results with suzetrigine, we characterized the effects of a selective Nav.7 inhibitor, AM-2099, on action potential generation and repetitive firing of dissociated human dorsal root ganglion neurons, studied at 37°C. Inhibition of Nav1.7 channels by 600 nM AM-2099 generally produced a substantial depolarizing shift of action potential threshold, an increase in rheobase, a decrease in action potential upstroke velocity, decrease in action potential peak, and prolongation of refractory period. Compared to inhibition of Nav1.8 channels, inhibition of Nav1.7 channels had larger effects on threshold and maximal upstroke velocity, while action potential peak was reduced similarly by both. Nav1.8 inhibition produced much more dramatic reduction of repetitive firing than Nav1.7 inhibition. The results show that although the excitability of human DRG neurons is affected by inhibition of Nav1.7 channels, most notably by an increase in threshold and increase in refractory period, repetitive firing of the neurons in response to strong stimuli is little affected.
Significance statement
Nav1.7 sodium channels are highly expressed in primary pain-sensing neurons and humans with null mutations in Nav1.7 channels have loss of pain sensation. However, unlike the Nav1.8 inhibitor suzetrigine, Nav1.7 inhibitors have so far not reached clinical use. We compared effects of Nav1.7 on electrical excitability of human dorsal root ganglion neurons with those of suzetrigine and found that while Nav1.7 inhibition affects spike threshold more than suzetrigine, there is little effect on repetitive firing with strong stimuli.
Introduction
The discovery of families with congenital insensitivity to pain traced to loss of function mutations in Nav1.7 voltage-dependent sodium channels (
Cox et al., 2006
;
Goldberg et al., 2007
; reviewed by
Dib-Hajj et al., 2013
;
Drissi et al., 2020
) suggested that Nav1.7 channels could be an ideal pharmacological target and launched many drug development efforts. However, PF-05089771, the first selective Nav1.7-inhibitor to reach large-scale clinical trials, was much less effective on pain in humans than anticipated (
McDonnell et al., 2018
;
Siebenga et al., 2020
; reviewed by
Alsaloum et al., 2020
) and many though not all Nav1.7-focused drug development efforts have been discontinued (
Eagles et al., 2022
;
Alsaloum et al., 2025
; Banh et al., 2025;
Yang et al., 2025
).
The reasons that Nav1.7 inhibitors have so far failed to advance to successful clinical studies are still unclear. One potential limitation of selectively inhibiting Nav1.7 channels is that primary pain-sensing neurons (nociceptors) express not only Nav1.7 channels but also have prominent expression of Nav1.8 channels (reviewed by
Cummins et al., 2007
;
Bennett et al. 2019
;
Alles et al., 2021
;
Goodwin and McMahon, 2021
). In the cell bodies of primary nociceptors of rat and mouse, Nav1.8 channels carry most of the sodium current driving the action potential
Renganathan et al., 2001
;
Blair and Bean, 2002
; reviewed by
Han et al., 2016
) and are particularly important for supporting repetitive firing of the neurons (
Elliott and Elliott, 1993
;
Cummins and Waxman, 1997
;
Rush et al., 1998
;
Renganathan et al., 2001
;
Blair and Bean, 2003
;
Matsutomi et al., 2006
;
Patrick Harty and Waxman, 2007
;
Han et al., 2015
). Recently, a potent and highly-selective Nav1.8 inhibitor, suzetrigine, was approved by the United States Food and Drug Administration for treatment of acute pain based on results in clinical studies on post-operative pain (
Jones et al., 2023
;
Bertoch et al., 2025
). However, although suzetrigine was more effective than placebo in these studies on post-operative pain, it was far from producing complete analgesia.
We lack a good understanding of the different roles of Nav1.7 and Nav1.8 channels in controlling the excitability of human primary nociceptors. Inhibitors of both Nav1.7 and Nav1.8 channels have been shown to reduce excitability of dorsal root ganglion neurons from human donors (
Payne et al., 2015
;
Alexandrou et al., 2016
;
Osteen et al., 2025
;
Stewart et al., 2025
), but for studies of Nav1.7 inhibitors, data is limited to a handful of cells studied with limited protocols (
Alexandrou et al., 2016
). Studies with suzetrigine inhibition of Nav1.8 channels showed only modest effects on action potential threshold and upstroke in most human DRG neurons (
Stewart et al., 2025
), suggesting a possible major role of Nav1.7 channels. Here we have examined effects of a Nav1.7 inhibitor, AM-2099, on excitability of DRG neurons from human neurons. We found that Nav1.7 inhibition has larger effects than Nav1.8 inhibition on action potential threshold and action potential upstroke but much less effect on ability of the neurons to fire repetitively during stimulation by maintained depolarization. Combined with the results of a similar study with suzetrigine, the results show that selective inhibition of either Nav1.7 or Nav1.8 channels alone has limited efficacy for inhibiting firing of human DRG neurons.
Results
We selected AM-2099 as a suitable Nav1.7 inhibitor, based on its potency and high degree of selectivity, >200-fold for Nav1.7 over Nav1.8 (
Marx et al., 2016
).
Figure 1
shows the effects of 600 nM AM-2099 on action potentials in human DRG neurons studied at 37°C. Action potentials were evoked by short (0.5-ms) current injections. With this protocol, the action potential shape is not affected by the stimulating current, making it easier to interpret changes in action potential shape produced by channel inhibition.
Figure 1A-B
shows a typical effect of AM-2099: a slowing of the rising phase of the action potential and substantially reduced peak, with little effect on the falling phase or after-hyperpolarization.
Figure 1C-E
show collected results for the effect of 600 nM AM-2099 on action potential parameters. In collected results, 600 nM AM-2099 produced a consistent decrease in maximum upstroke velocity to an average of 42 ± 6% of control, from 337 ± 42 to 154 ± 30 mV/mS (mean ± SEM, n=18; p=.0002, two-tailed Wilcoxon test), and a consistent decrease in action potential peak by an average of 16 ± 3 mV, from +30 ± 3 mV to +14 ± 5 mV (n=18; p=.0002, two-tailed test Wilcoxon test). Unlike the substantial decrease in the width of the action potential seen with suzetrigine inhibition of Nav1.8 channels (
Stewart et al., 2025
), inhibiting Nav1.7 channels by AM-2099 did not have a consistent effect on action potential width, increasing the width in 5 of 13 neurons and decreasing the width in 8 neurons (p =0.2, two-tailed test Wilcoxon test; width measured at 0 mV, with results including only neurons in which the peak was > 0 mV n=13).
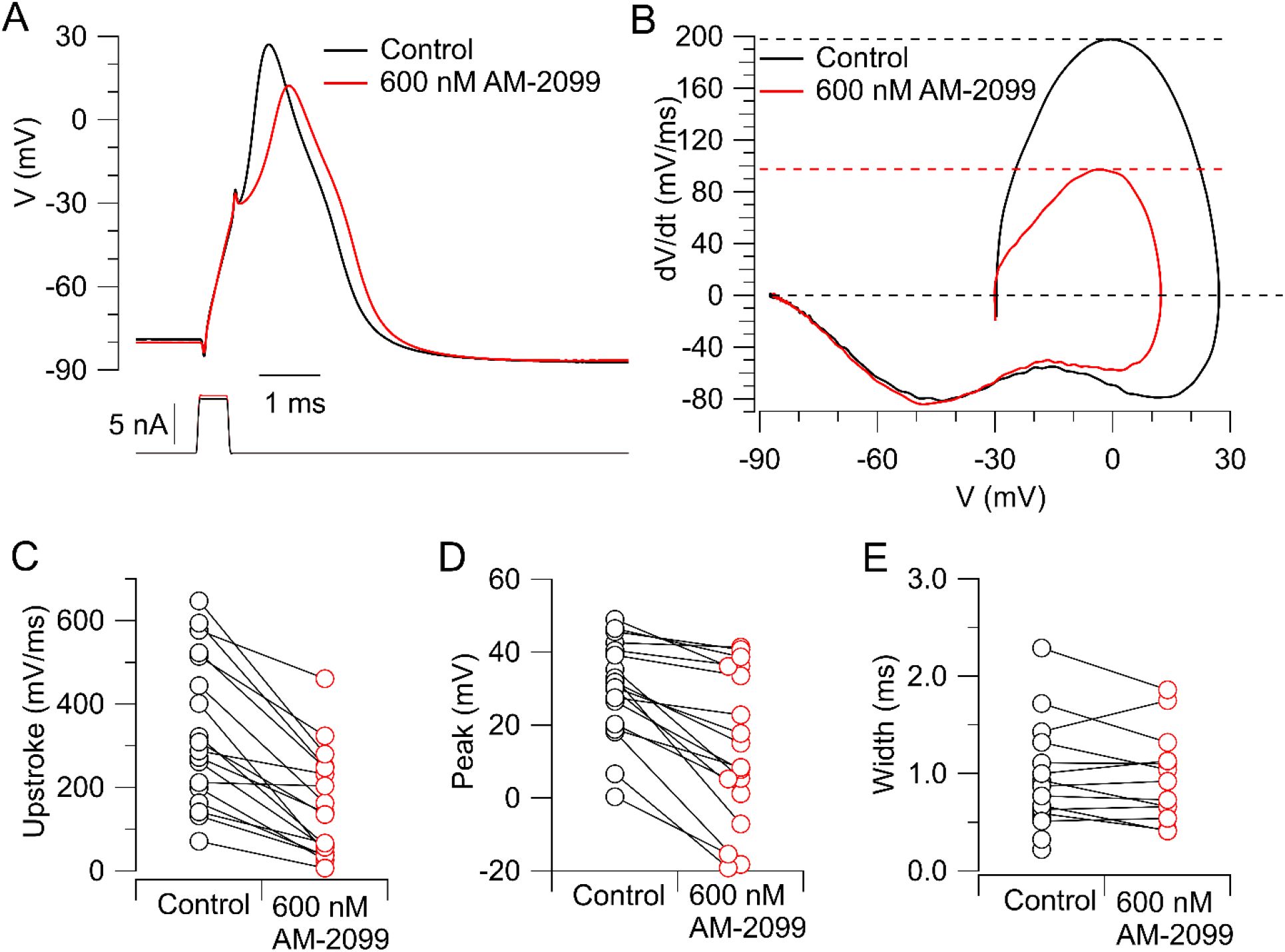
Effect of AM-2099 on action potentials in human DRG neurons.
A, Action potentials evoked by a short (0.5-ms) current injection before and after application of 600 nM AM-2099. Stimulating current was adjusted to be slightly larger in AM-2099 so that the voltage immediately after cessation of the stimulating current was the same as in control (because control has an active component in the last 0.1 ms of the stimulating pulse that is inhibited with AM-2099). B, Phase-plane plot of dV/dt versus V showing reduction of maximum upstroke velocity and peak. Upper dashed lines: maximum upstroke velocity in control (black) and with 600 nM AM-2099 (red). C, Collected results for effect of 600 nM AM-2099 on the maximal upstroke velocity of the action potential in 18 neurons. D, Collected results for effect of 600 nM AM-2099 on peak of the action potential evoked by a short current injection. E, Collected results for effect of 600 nM AM-2099 on the width of the action potential measured at 0 mV. In cells with values only in control, the action potential in AM-2099 had a peak < 0 mV.
AM-2099 consistently produced a depolarizing shift in the action potential threshold, determined by short (0.5-ms) current injections of increasing magnitude (
Figure 2
). In collected results, the threshold increased by an average of 9.4 ± 1.2 mV from -44.7 ± 2.2 mV to -35.4 ± 3.0 mV (n=18; p=.0002, two-tailed test Wilcoxon test). The 0.5-ms current injection required to reach threshold increased by an average of 20 ± 3% (n=18; p=.0002, two-tailed test Wilcoxon test).
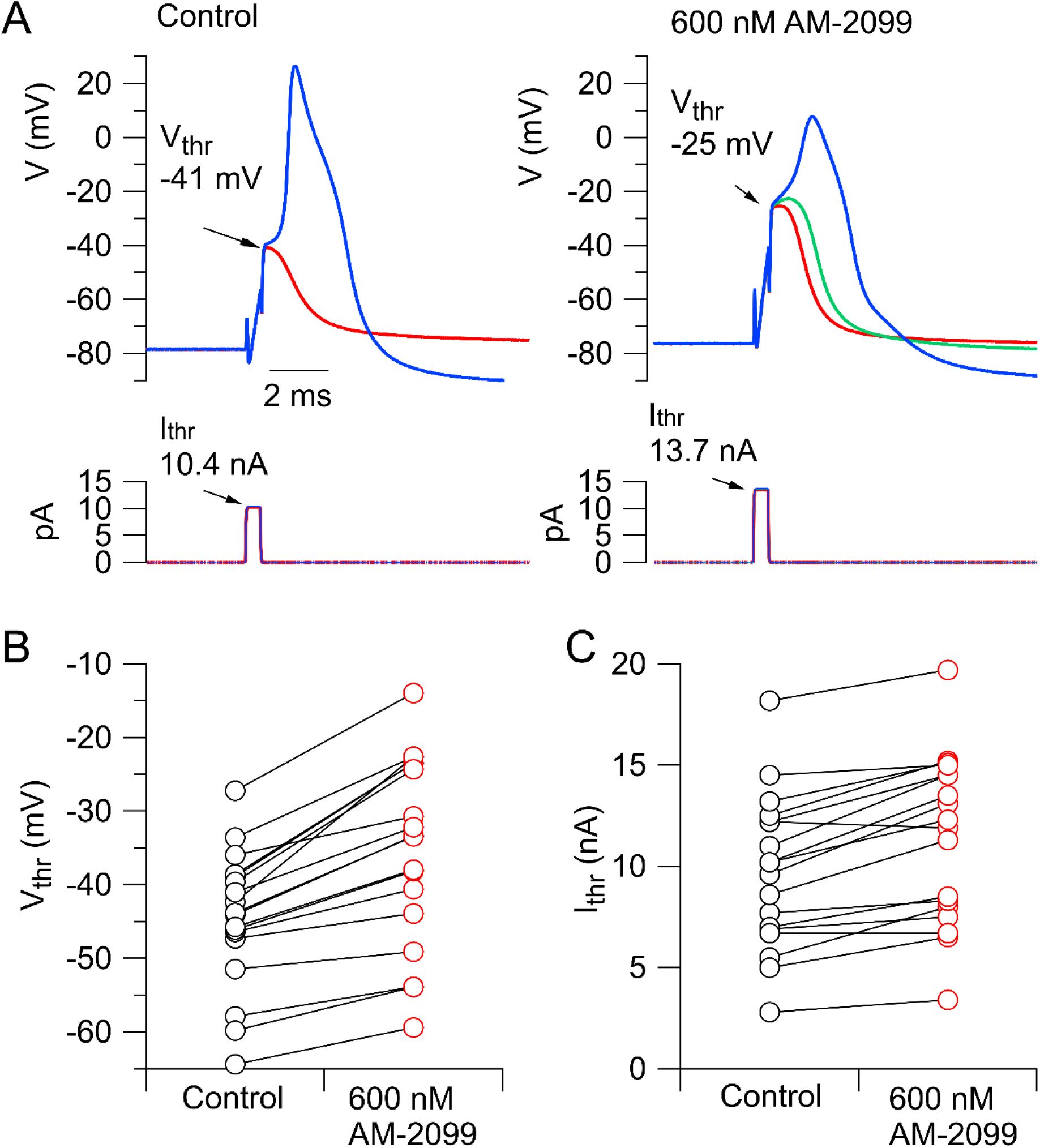
Effect of AM-2099 on action potential threshold.
Short (0.5-ms) current injections of different size were delivered, with gradations to find the level of current injection that first evoked an action potential. The voltage immediately after the smallest current injection that evoked a spike was considered as the threshold voltage. A, Just sub-threshold and supra-threshold current injections in control (left) and after application of 600 nM AM-2099 (right). B, Collected data for threshold voltage before and after 600 AM-2099 (n=18). C, Collected data for minimum current (0.5-ms current injection) that evoked an action potential (n=18).
We next explored the effect of inhibiting Nav1.7 channels on repetitive firing evoked by 1-s current injections.
Figure 3A-B
shows a typical example: the minimal current required to elicit a spike (rheobase) increased substantially but the frequency of firing evoked by large current injections did not change much. In collected results (
Figure 3C
), rheobase current increased by 69 ± 12%, from 2.4 ± 0.3 nA to 4.1 ± 0.6 nA (mean ± SEM, n=23; p=.0002, two-tailed Wilcoxon test). In 25 neurons tested, 14 fired multiple action potentials in control, and 13 of the 14 still fired multiple action potentials after application of AM-2099 (
Figure 3D
).
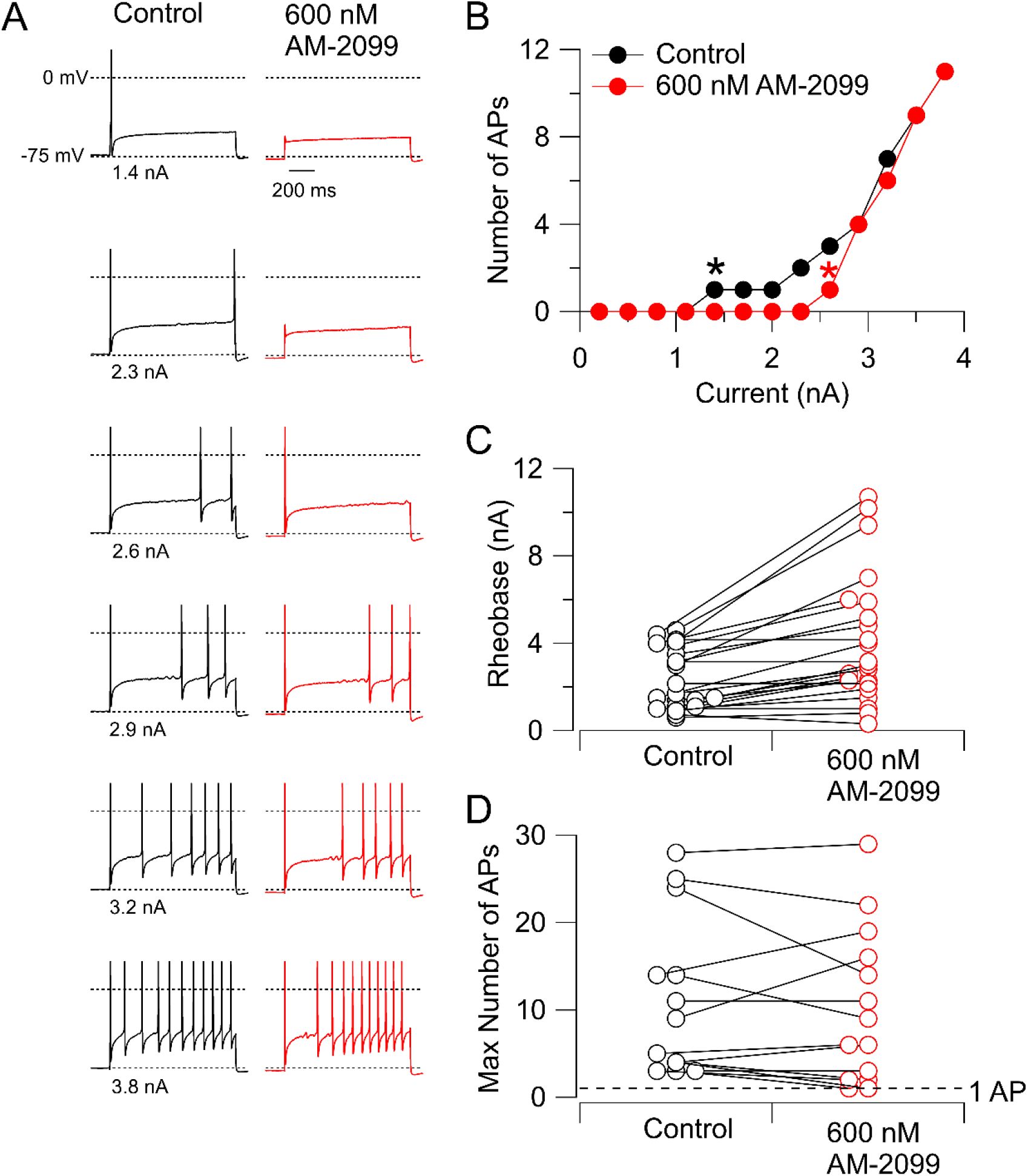
Effect of 600 nM AM-2099 on rheobase and repetitive firing.
A, Firing evoked by 1-s injections of current of increasing magnitude in control and after application of 600 nM AM-2099. B, Number of action potentials as a function of the injected current before and after 600 nM AM-2099 in this neuron. Asterisks indicate rheobase current in control (black) and in AM-2099 (red). C, Collected data for the effect of 600 nM AM-2099 on rheobase current. D, Collected results for the effect of 600 nM AM-2099 on the maximal number of action potentials during 1-s current injections over a range of magnitudes for neurons that fired more than one action potential in control. Dashed line drawn at 1 action potential.
In a previous study, we were surprised to find that inhibiting Nav1.8 channels with suzetrigine had the counter-intuitive effect of decreasing the refractory period (
Stewart et al., 2025
). In contrast, inhibiting Nav1.7 channels resulted in an increase in the refractory period in 14 of 15 neurons tested (
Figure 4
), with an average increase to 1.8 ± 0.2 of the value in the control (mean ± SEM, n=15; p=.0013, two-tailed Wilcoxon test).
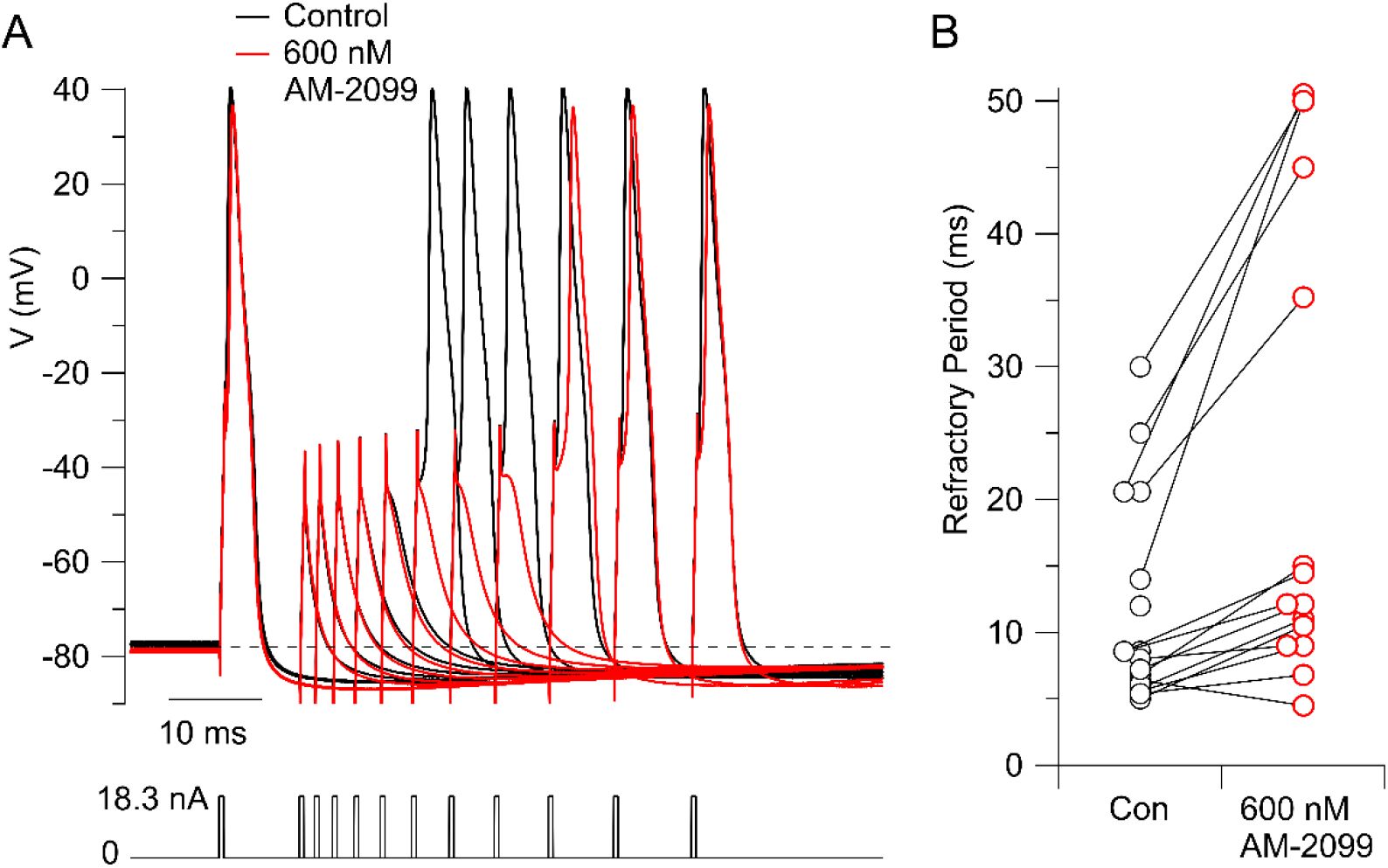
Prolongation of refractory period by AM-2099.
A, Action potentials were evoked by a pair of 0.5-ms current injections with a variable time between them, with magnitude of both set at 1.5-times the threshold current determined in control. The time between the two current injections was varied from longer to shorter to determine the refractory period with this stimulus. The figure shows superimposed sweeps from 11 different sets of times in control (black) and with 600 nm AM-2099 (red). In control, the second stimulus evoked an action potential with a spacing (start to start) of 20.6 ms, while after AM-2099, a spacing of 35.2 ms was required to evoke an action potential using the same stimuli. B, Collected results in 15 neurons.
Discussion
Previous work has shown that, like mouse and rat small-diameter dorsal root ganglion neurons (reviewed by
Rush et al., 2007
;
Bennett et al. 2019
;
Goodwin and McMahon, 2021
), human dorsal root ganglion neurons express both Nav1.7 and Nav1.8 channels (
Payne et al., 2015
;
Alexandrou et al., 2016
;
Zhang et al., 2017
;
Osteen et al., 2025
). In a previous paper, we used the selective Nav1.8 inhibitor suzetrigine (VX-548) to explore how inhibition of Nav1.8 channels affects action potential generation in the neurons (
Stewart et al., 2025
). The results presented here using similar protocols show that inhibiting Nav1.7 channels also strongly affects the excitability of human DRG neurons but in different ways than inhibiting Nav1.8 channels.
Figure 5
compares the effects of inhibiting Nav1.7 channels on action potential threshold, shape, and ability to fire repetitively with the effects of Nav1.8 inhibition determined in a previous study (
Stewart et al., 2025
), with recordings in both cases done at 37°C with the same protocols. Inhibiting Nav1.7 channels generally produces a larger effect on action potential threshold (
Figure 5A
) and maximum upstroke (
Figure 5B
) than Nav1.8 inhibition, while the effects on action potential peak were similar (
Figure 5C
). A major difference is that inhibiting Nav1.8 channels was generally much more effective than Nav1.7 inhibition in reducing repetitive firing during 1-sec current injections (
Figure 5D
).
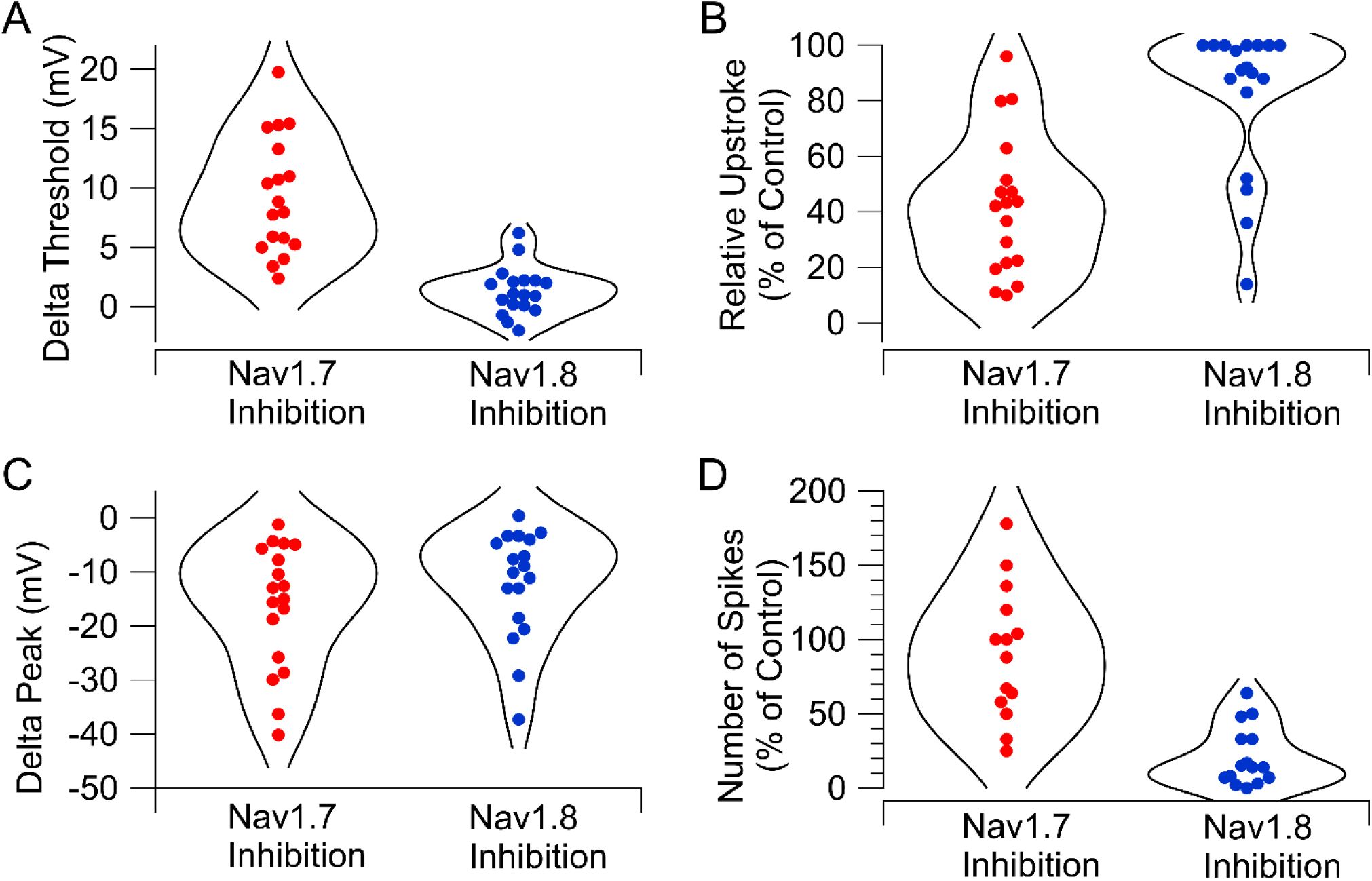
Comparison of Nav1.7 and Nav1.8 inhibition on action potential firing in human DRG neurons.
A, Change in action potential threshold by inhibiting Nav1.7 channels (600 nM AM-2099) or Nav1.8 channels (10 nM VX-548). B, Inhibition of maximum upstroke of action potential by inhibiting Nav1.7 channels (600 nM AM-2099) or Nav1.8 channels (10 nM VX-548). C, Reduction of action potential peak by inhibiting Nav1.7 channels (600 nM AM-2099) or Nav1.8 channels (10 nM VX-548). D, Effect of inhibiting Nav1.7 channels (600 nM AM-2099) or Nav1.8 channels (100 nM VX-548) on maximum number of action potentials evoked by 1-s depolarizations of different magnitudes. Data for effects of 100 nM VX-548 from
Stewart et al., 2025
.
The different effects of inhibiting Nav1.7 and Nav1.8 channels on action potential shape and repetitive firing fit well with their different voltage-dependence and kinetics as defined from studies on both human and rodent channels (
Alexandrou et al., 2016
;
Köster et al., 2025
;
Vasylyev et al., 2025a,b
) and with the different but overlapping roles of Nav1.7 and Nav1.8 channels in action potentials of mouse and rat DRG neurons (
Vasylyev et al., 2025a,b
;
Xie et al., 2024
). Nav1.7 channels are activated more quickly and at more negative voltages than Nav1.8 channels, consistent with the dominant effect of Nav1.7 channels on threshold and maximum upstroke velocity. Nav1.8 channels inactivate more slowly than Nav1.7 channels and a substantial fraction of Nav1.8 channels remain open during the shoulder of the action potential (
Stewart et al., 2025
). An apparent quantitative difference between human and rodent DRG neurons is the dominance of Nav1.7 channels in the upstroke of the action potential in human neurons, while TTX-resistent current from Nav1.8 channels appears to dominate in the upstroke of action potentials in small-diameter rat and mouse neurons (
Renganathan et al., 2001
;
Blair and Bean, 2002
;
Matsutomi et al., 2006
;
Rush et al., 2007
;
Han et al., 2015a
). Interestingly, this quantitative difference between mouse and rat DRG neurons was mirrored in a PCR analysis of seven sodium channel subtypes in DRG neurons of human and mouse, showing higher relative expression of Nav1.7 compared to Nav1.8 in human neurons compared to mouse neurons (
Chang et al., 2018
).
Our results are generally consistent with those of
Alexandrou et al. (2016)
, who found that the Nav1.7 inhibitor PF-05089771 blocked action potential generation in most but not all (8 of 15) human DRG neurons tested when action potentials were evoked by current injections that were just-suprathreshold in control. We saw more consistent effects of inhibiting Nav1.7 channels, with an increase in voltage threshold in 18 of 18 neurons. The difference could reflect in part the different recording conditions (37°C in our experiments versus room temperature in the Alexandrou et al. recordings) but could also reflect intrinsic heterogeneity in cell populations.
The strong effect of inhibiting Nav1.7 current on action potential threshold is reflected in the increase in rheobase current necessary to evoke spiking in the 1-s current injection protocols (
Figure 3
). The effect of inhibiting Nav1.7 channels with AM-2099 is concordant with an increase in rheobase in iPSC-derived nociceptors from human patients with Nav1.7 loss-of-function mutations (
McDermott et al., 2019
) and with an increase in rheobase with the Nav1.7 inhibitor PF-05089771 in iPSC neur
McDermott et al., 2019
ons from healthy controls but not patients with Nav1.7 loss-of-function mutations (
McDermott et al., 2019
).
In contrast to the counter-intuitive decrease in refractory period seen with Nav1.8 inhibition (
Stewart et al., 2025
), Nav1.7 inhibition almost always (14 of 15 neurons) increased the refractory period. The overall effects on refractory period of inhibiting either channel reflect a complex combination of the actual reduction in sodium channel availability along with secondary effects resulting from the change in action potential shape. The increase in refractory period with Nav1.7 inhibition is easily understood from the fact that Nav1.7 channels strongly control action potential threshold. The reduction in action potential peak (seen with both Nav1.7 and Nav1.8 inhibition) and a reduction in action potential width (which was dramatic with Nav1.8 inhibition but generally not with Nav1.7 inhibition) would both be expected to have a secondary effect of tending to reduce the refractory period by producing less activation of voltage-activated potassium channels and therefore reducing the post-spike potassium conductance that contributes to the refractory period. Because inhibiting Nav1.8 channels has much less effect on action potential threshold compared to inhibiting Nav1.7 channels, the effects on potassium conductance to decrease refractory period – resulting from the changes in action potential shape - apparently dominate with Nav1.8 inhibition but not Nav1.7 inhibition.
A major limitation in our current knowledge of nociceptor excitability is that while we have relatively detailed information on ion channel composition and electrophysiological properties of cell bodies, we have only limited information about the electrophysiology of the key functional elements of the neurons: the peripheral terminals where action potentials are first generated, the axons where the action potentials are propagated, and the fine branches and synaptic boutons of the central terminals where action potentials trigger synaptic transmission (
Bennett et al., 2019
;
Goodwin and McMahon, 2021
). Nav1.7 channels are clearly involved in generation and propagation of action potentials, because selective Nav1.7 inhibitors applied to the nerve can inhibit firing of C-fibers evoked by cutaneous stimulation, although often incompletely (
Goodwin et al., 2022
;
Deng et al., 2023
). A recent study making
in vivo
pig single nerve fiber recordings of C-fiber nociceptors excited by transcutaneous stimulation found that inhibiting Nav1.7 channels by protoxin II reduced firing more dramatically at low stimulus intensities than high intensities (
Soares et al., 2025
), which could be consistent with Nav1.7 channels regulating firing near threshold stimulation but Nav1.8 channels being more important for repetitive firing with strong stimulation. In general, although studies on the cell body probably have major limitations for predicting clinical effects, they can help define the pharmacological properties of individual channels and perhaps elucidate some ways that overall firing patterns can be regulated by particular channels. For example, the different effects on refractory period and on repetitive firing of inhibiting Nav1.8 versus Nav1.7 channels seems likely to apply wherever both are expressed.
Despite the clear differences between Nav1.7 and Nav1.8 channel inhibition across the entire population of cells tested, there is striking cell-to-cell variability in the effects of inhibiting either Nav1.7 or Nav1.8 channels. For example, although AM-2099 generally had small effects on maximal repetitive firing, it reduced repetitive firing to half or less of control values in 3 of the 14 neurons that had repetitive firing in control (
Figures 3
,
5D
). The relatively large effects of Nav1.7 inhibition on repetitive firing in a subset of neurons is consistent with a previous study quantifying components of sodium current using action potential clamp, in which 2 of 10 neurons tested had a large contribution of Nav1.7 current in the second action potential during repetitive spiking (
Stewart et al., 2025
). Similarly, although VX-548 inhibition of Nav1.8 channels had only a small effect on maximal upstroke velocity in most cells, in 4 of 18 cells upstroke velocity was reduced to less than 50% of the control value, similar to the typical effect of Nav1.7 inhibition. In the future, larger data sets in combination with single cell measurements of mRNA levels may help interpret the large cell-to-cell variability in terms of defined subpopulations of C-fiber nociceptors (cf.
Shiers et al., 2020
;
Middleton et al., 2021
;
Tavares-Ferreira et al., 2022
;
Körner et al., 2022
;
2026
).
Clinical studies with the first selective Nav1.7 inhibitors to reach studies on human pain showed relatively poor efficacy compared to the Nav1.8 inhibitor suzetrigine (reviewed by
Eagles et al., 2022
;
Alsaloum et al., 2025
;
Yang et al., 2025
). The reasons for the low efficacy of the Nav1.7 inhibitors so far tested on human pain are unclear, and may include poor target engagement as a result of high plasma protein binding (
Deng et al., 2023
). Recognizing the limitations of studies on cell bodies, our results raise the possibility that another factor distinguishing the two targets could be the greater effect of Nav1.8 inhibition to reduce repetitive firing of the neurons compared to Nav1.7 inhibition. It seems plausible that this difference may extend to initial generation of neuronal firing in the peripheral terminals, where computer modeling suggests that Nav1.8 channels play a major role in regulating repetitive firing (
Barkai et al., 2020
). The perception of pain is correlated with higher firing frequencies of nociceptors (
Yarnitsky et al., 1992
;
Djouhri et al., 2006
; reviewed by
Namer and Lampert, 2025
), so it is possible that the greater efficacy of Nav1.8 inhibition compared to Nav1.7 inhibition to reduce repetitive firing of human nociceptive neurons contributes to differences in clinical efficacy.
The expression of Nav1.7 channels can be modified in various pathological conditions associated with pain, including as chemotherapy-induced peripheral neuropathy (
Li et al., 2018
;
Akin et al., 2021
;
Braden et al., 2022
) and early-life stress (
Alvarez et al., 2021
). The efficacy of various Nav1.7 inhibitors in a variety of animal models of pain suggests that there may be particular forms of human pain that would respond to Nav1.7 inhibition more robustly than the acute pain from initially tested. Further studies of roles of Nav1.7 channels in excitability of human nociceptors in conditions such as chemotherapy-induced peripheral neuropathy and other forms of human neuropathic pain (
Raja et al., 2020
;
Stucky and Mikesell, 2021
) would be illuminating and may be feasible as it becomes possible to test neurons from human donors with known pain conditions.
Materials and Methods
Preparation of human DRG neurons
Neurons were obtained from the dorsal root ganglia (DRGs) of human donors. The procurement network of AnaBios Corporation includes only US-based Organ Procurement Organizations and Hospitals. Policies for donor screening and consent are those established by the United Network for Organ Sharing (UNOS). Organizations supplying human tissues to AnaBios follow the standards and procedures established by the US Centers for Disease Control (CDC) and are inspected biannually by the Department of Health and Human Services (DHHS). The distribution of donor medical information is in compliance with HIPAA regulations to protect donor privacy. All transfers of donor tissue to AnaBios are fully traceable and periodically reviewed by US Federal authorities.
Neurons were dissociated from dorsal root ganglia and suspended in medium (
Davidson et al., 2014
) and shipped in suspension in a container with temperature maintained at 4°C (
Lesnak et al., 2025
). The neurons were then plated on round 12 mm poly-D-lysine-treated coverslips placed in 24- or 48-well plates. Coverslips (Fisherbrand, Cat#12-545-80) were prepared by exposure UV for approximately 15-30 minutes and then incubated overnight at 4°C with 0.01 mg/mL poly-D-lysine diluted in sterile water. After trituration of the tissue, 60 μL of the sample was plated on each coverslip and incubated at 37°C (5% CO
2
) for 1-2 hours to allow the cells to settle and attach to the coverslip. Each well was then gently flooded with 1 mL of culture media consisting of BrainPhys media (Stemcell technologies, Cat. 05709), 1% penicillin/streptomycin, 1% GlutaMAX, 2% NeuroCult SM1 (Stemcell technologies, Cat 05711), 1% N-2 Supplement (Thermo Scientific, Cat # 17502048) and 2% Fetal Bovine Serum. Plates were housed in a 5% CO
2
incubator set at 37°C for up to 6 days.
Electrophysiology with human DRG neurons
For recording, coverslips were placed into the recording chamber containing about 2 mL of Tyrode’s solution at room temperature. We generally selected smaller cells for recording and selected cells in which glia had partially peeled off, which facilitated recording. Prior to recording, if the cell was strongly adhered to the coverslip (as was usual 2-3 days after plating), an electrode with a blunt tip was used to scrape the surrounding area of the cell and gently maneuvered to ensure detachment from the coverslip. Both the “scraping” electrode and recording electrode were pulled from borosilicate capillaries (VWR International, Cat #53432-921) on a Sutter P-97 puller (Sutter Instruments).
Whole-cell patch-clamp recordings were made with either an Axon Instruments Multiclamp 700B amplifier (Molecular Devices) controlled by pClamp9.2 software (Axon Instruments), filtered at 10 kHz with a low-pass Bessel filter, and digitized at 100 kHz by a Digidata 1322A data acquisition interface or a Sutter Instruments dPatch integrated amplifier/data acquisition system, with filtering at 10 kHz and digitization at 100 kHz. Whole-cell recordings were obtained using patch pipettes with resistances of 1.2-2.5 MΩ. The shank of the electrode was wrapped with Parafilm (American National Can Company) in order to reduce capacitance and allow optimal series resistance compensation (bridge balance) of 60-80% without oscillation. Seals were obtained and the whole-cell configuration established with cells in room-temperature Tyrode’s solution consisting of 155 mM NaCl, 3.5 mM KCl, 1.5 mM CaCl
2
, 1 mM MgCl
2
, 10 mM HEPES, 10 mM glucose, pH adjusted to 7.4 with NaOH. After establishing whole-cell recording, cells were lifted off the bottom of the recording chamber and placed in front of an array of quartz flow pipes (250 μm internal diameter, 350 μm external diameter, Polymicro Technologies) attached with styrene butadiene glue (Amazing Goop, Eclectic Products) to an aluminum rod (1×1 cm) whose temperature was controlled by resistive heating elements and a feedback-controlled temperature controller (TC-344B; Warner Instruments). Solution changes were made (in < 1 second) by moving the cell between adjacent pipes. Experiments were done with temperature controlled at 37°C. Whole-cell recordings were obtained using patch pipettes filled with a K-gluconate-based internal solution containing (in mM) 139.5 K-Gluconate, 1.6 MgCl
2
, 1 EGTA, 0.09 CaCl
2
, 9 HEPES,14 creatine phosphate (Tris salt), 4 MgATP, 0.3 GTP (Tris salt), pH adjusted to 7.2 with KOH. Membrane potentials are corrected for a liquid junction potential of -13 mV between the internal solution and the Tyrode’s solution in which the current was zeroed before recording.
Cells were recorded at their natural resting membrane potential without any injection of steady holding current. For the data in
Figures 1
,
2
, and
4
, action potentials were evoked by short (0.5-ms) injections of current so that the action potential occurred after the current injection. For the data in
Figure 3
,
1-s
current steps were applied with the increments of the steps adjusted depending on the input resistance of each cell. Action potentials were defined using a criteria of action potential peak >-20 mV and height >40 mV. Experiments were included in combined data sets only if the resting potential varied by less than 5 mV during the application of AM-2099. Capsaicin sensitivity was tested at the end of the experiment by holding the cell at -70 mV in voltage clamp and briefly applying 1 μM capsaicin.
Drugs
AM-2099 (
Marx et al., 2016
) was purchased from MedChemExpress (Catalog HY-100727). A stock solution was prepared at 10 mM in DMSO, aliquoted, and frozen at - 20°C. Capsaicin stock was prepared at 1 mM from powder in DMSO and stored at room temperature. External control solutions contained concentrations of DMSO to match those in the drug-containing solutions. All external solutions contained 1 mg/mL Pluronic PF-68 (Sigma).
Data analysis and statistics
Data were analyzed using programs written in Igor Pro 6, 8 or 9 (Wavemetrics, Lake Oswego, OR), using DataAccess (Bruxton Software) to read pClamp files into Igor Pro.
Data availability
All data is contained in figures.
Acknowledgements
This work was supported by National Institutes of Health Grant R35-NS127216. Funds supporting acquisition and preparation of the neurons were from the AnaBios Coporation. We are very grateful to the anonymous donors and their families for providing the human DRG neurons used in this study. We thank Richard Kondo for facilitating transfer of the preparation of the human DRG tissue.
Additional information
Author contributions
AF, SJ, RGS, and TO designed and executed experiments, analyzed data, and contributed to writing the manuscript; AF and KC designed procedures for and supervised dissection and preparation of tissue; BPB helped design experiments, analyze data, and write the manuscript.
Funding
HHS | NIH | National Institute of Neurological Disorders and Stroke (NINDS) (R35-NS127216)
Bruce P Bean
References
Paclitaxel increases axonal localization and vesicular trafficking of Nav1.7
Brain
144
:1727–1737
PubMed
Google Scholar
Subtype-Selective Small Molecule Inhibitors Reveal a Fundamental Role for Nav1.7 in Nociceptor Electrogenesis, Axonal Conduction and Presynaptic Release
PLoS One
11
:e0152405
PubMed
Google Scholar
Pharmacological inhibition of NaV1.8 by suzetrigine reveals potent analgesic potential without tolerance development in mice
Mol Brain
18
:86
PubMed
Google Scholar
Peripheral Voltage-Gated Cation Channels in Neuropathic Pain and Their Potential as Therapeutic Targets
Front Pain Res (Lausanne)
2
:750583
PubMed
Google Scholar
Voltage-gated sodium channels in excitable cells as drug targets
Nat Rev Drug Discov
PubMed
Google Scholar
Status of peripheral sodium channel blockers for non-addictive pain treatment
Nat Rev Neurol
16
:689–705
PubMed
Google Scholar
Nociceptor Overexpression of Na(V)1.7 Contributes to Chronic Muscle Pain Induced by Early-Life Stress
J Pain
22
:806–816
PubMed
Google Scholar
Sodium Channel Inhibitors in Clinical Development for Pain Management: A Focused Review
CNS Drugs
40
:165–180
PubMed
Google Scholar
The Input-Output Relation of Primary Nociceptive Neurons is Determined by the Morphology of the Peripheral Nociceptive Terminals
J Neurosci
40
:9346–9363
PubMed
Google Scholar
The Role of Voltage-Gated Sodium Channels in Pain Signaling
Physiol Rev
99
:1079–1151
PubMed
Google Scholar
Suzetrigine, a Nonopioid Na V 1.8 Inhibitor for Treatment of Moderate-to-severe Acute Pain: Two Phase 3 Randomized Clinical Trials
Anesthesiology
142
:1085–1099
PubMed
Google Scholar
Roles of tetrodotoxin (TTX)-sensitive Na+ current TTX-resistant Na+ current and Ca2+ current in the action potentials of nociceptive sensory neurons
J Neurosci
22
:10277–10290
PubMed
Google Scholar
Role of tetrodotoxin-resistant Na+ current slow inactivation in adaptation of action potential firing in small-diameter dorsal root ganglion neurons
J Neurosci
23
:10338–10350
PubMed
Google Scholar
Small molecule targeting NaV1.7 via inhibition of the CRMP2-Ubc9 interaction reduces and prevents pain chronification in a mouse model of oxaliplatin-induced neuropathic pain
Neurobiol Pain
11
:100082
PubMed
Google Scholar
Expression and Role of Voltage-Gated Sodium Channels in Human Dorsal Root Ganglion Neurons with Special Focus on Nav1.7, Species Differences, and Regulation by Paclitaxel
Neurosci Bull
34
:4–12
PubMed
Google Scholar
An SCN9A channelopathy causes congenital inability to experience pain
Nature
444
:894–898
PubMed
Google Scholar
The roles of sodium channels in nociception: Implications for mechanisms of pain
Pain
131
:243–257
PubMed
Google Scholar
Downregulation of tetrodotoxin-resistant sodium currents and upregulation of a rapidly repriming tetrodotoxin-sensitive sodium current in small spinal sensory neurons after nerve injury
J Neurosci
17
:3503–3514
PubMed
Google Scholar
Human sensory neurons: Membrane properties and sensitization by inflammatory mediators
Pain
155
:1861–1870
PubMed
Google Scholar
Nav1.7 is essential for nociceptor action potentials in the mouse in a manner independent of endogenous opioids
Neuron
111
:2642–2659
PubMed
Google Scholar
The Na(V)1.7 sodium channel: from molecule to man
Nature reviewsNeuroscience
14
:49–62
PubMed
Google Scholar
Association of somatic action potential shape with sensory receptive properties in guinea-pig dorsal root ganglion neurones
The Journal of physiology
513
:857–872
PubMed
Google Scholar
The TTX-resistant sodium channel Nav1.8 (SNS/PN3): expression and correlation with membrane properties in rat nociceptive primary afferent neurons
J Physiol
550
:739–752
PubMed
Google Scholar
Spontaneous pain, both neuropathic and inflammatory, is related to frequency of spontaneous firing in intact C-fiber nociceptors
J Neurosci
26
:1281–1292
PubMed
Google Scholar
Understanding the genetic basis of congenital insensitivity to pain
Br Med Bull
133
:65–78
PubMed
Google Scholar
Fifteen years of Na(V) 1.7 channels as an analgesic target: Why has excellent in vitro pharmacology not translated into in vivo analgesic efficacy?
Br J Pharmacol
179
:3592–3611
PubMed
Google Scholar
Characterization of TTX-sensitive and TTX-resistant sodium currents in small cells from adult rat dorsal root ganglia
J Physiol
463
:39–56
PubMed
Google Scholar
Global Nav1.7 knockout mice recapitulate the phenotype of human congenital indifference to pain
PLoS One
9
:e105895
PubMed
Google Scholar
Loss-of-function mutations in the Nav1.7 gene underlie congenital indifference to pain in multiple human populations
Clin Genet
71
:311–319
PubMed
Google Scholar
Identification and targeting of a unique Na(V)1.7 domain driving chronic pain
Proc Natl Acad Sci U S A
120
:e2217800120
PubMed
Google Scholar
The physiological function of different voltage-gated sodium channels in pain
Nat Rev Neurosci
22
:263–274
PubMed
Google Scholar
Examination of the contribution of Nav1.7 to axonal propagation in nociceptors
Pain
163
:e869–e881
PubMed
Google Scholar
Rat Na(V)1.7 loss-of-function genetic model: Deficient nociceptive and neuropathic pain behavior with retained olfactory function and intra-epidermal nerve fibers
Mol Pain
15
:1744806919881846
PubMed
Google Scholar
Human Na(v)1.8: enhanced persistent and ramp currents contribute to distinct firing properties of human DRG neurons
J Neurophysiol
113
:3172–3185
PubMed
Google Scholar
Sodium channel Nav1.8: Emerging links to human disease
Neurology
86
:473–483
PubMed
Google Scholar
Selective Inhibition of Na(V)1.8 with VX-548 for Acute Pain
N Engl J Med
389
:393–405
PubMed
Google Scholar
Sodium Channel Na(v)1.8 Underlies TTX-Resistant Axonal Action Potential Conduction in Somatosensory C-Fibers of Distal Cutaneous Nerves
J Neurosci
37
:5204–5214
PubMed
Google Scholar
Molecular architecture of human dermal sleeping nociceptors
Cell
:S0092-8674(25)01497-7
PubMed
Google Scholar
Functional subgroups of rat and human sensory neurons: a systematic review of electrophysiological properties
Pflugers Arch
474
:367–385
PubMed
Google Scholar
Nociceptor sodium channels shape subthreshold phase, upstroke, and shoulder of action potentials
J Gen Physiol
157
PubMed
Google Scholar
Enabling wider access to human molecular neuroscience research in pain: A simple preservation method for human dorsal root ganglion neurons in Hibernate A media
bioRxiv
:2648
PubMed
Google Scholar
DRG Voltage-Gated Sodium Channel 1.7 Is Upregulated in Paclitaxel-Induced Neuropathy in Rats and in Humans with Neuropathic Pain
J Neurosci
38
:1124–1136
PubMed
Google Scholar
Sulfonamides as Selective Na(V)1.7 Inhibitors: Optimizing Potency and Pharmacokinetics to Enable in Vivo Target Engagement
ACS Med Chem Lett
7
:1062–1067
PubMed
Google Scholar
Multiple types of Na(+) currents mediate action potential electrogenesis in small neurons of mouse dorsal root ganglia
Pflugers Arch
453
:83–96
PubMed
Google Scholar
Defining the Functional Role of Na(V)1.7 in Human Nociception
Neuron
101
:905–919
PubMed
Google Scholar
Efficacy of the Nav1.7 blocker PF-05089771 in a randomised, placebo-controlled, double-blind clinical study in subjects with painful diabetic peripheral neuropathy
Pain
159
:1465–1476
PubMed
Google Scholar
New Author Guidelines for Displaying Data and Reporting Data Analysis and Statistical Methods in Experimental Biology
Mol Pharmacol
97
:49–60
PubMed
Google Scholar
Nav1.7 is required for normal C-low threshold mechanoreceptor function in humans and mice
Brain
145
:3637–3653
PubMed
Google Scholar
Studying human nociceptors: from fundamentals to clinic
Brain
144
:1312–1335
PubMed
Google Scholar
Functional signatures of human somatosensory C fibers by microneurography
Pain
166
:2220–2235
PubMed
Google Scholar
Chemical Synthesis, Proper Folding, Na(v) Channel Selectivity Profile and Analgesic Properties of the Spider Peptide Phlotoxin 1
Toxins (Basel)
11
PubMed
Google Scholar
Pharmacology and Mechanism of Action of Suzetrigine, a Potent and Selective Na(V)1.8 Pain Signal Inhibitor for the Treatment of Moderate to Severe Pain
Pain Ther
PubMed
Google Scholar
Inactivation properties of sodium channel Nav1.8 maintain action potential amplitude in small DRG neurons in the context of depolarization
Mol Pain
3
:12
PubMed
Google Scholar
A novel selective and orally bioavailable Nav 1.8 channel blocker, PF-01247324, attenuates nociception and sensory neuron excitability
Br J Pharmacol
172
:2654–2670
PubMed
Google Scholar
John J Bonica Award Lecture: Peripheral neuronal hyperexcitability: the “low-hanging” target for safe therapeutic strategies in neuropathic pain
Pain
161
:S14–S26
PubMed
Google Scholar
EXPRESS: Voltage-dependent sodium (NaV) channels in group IV sensory afferents
Mol Pain
12
PubMed
Google Scholar
Contribution of Na(v)1.8 sodium channels to action potential electrogenesis in DRG neurons
J Neurophysiol
86
:629–640
PubMed
Google Scholar
Differential properties of tetrodotoxin-sensitive and tetrodotoxin-resistant sodium channels in rat dorsal root ganglion neurons
J Neurosci
12
:2104–2111
PubMed
Google Scholar
Electrophysiological characterization of ectopic spontaneous discharge in axotomized and intact fibers upon nerve transection: a role in spontaneous pain?
Pflugers Arch
474
:387–396
PubMed
Google Scholar
Electrophysiological properties of sodium current subtypes in small cells from adult rat dorsal root ganglia
J Physiol
511
:771–789
PubMed
Google Scholar
Multiple sodium channels and their roles in electrogenesis within dorsal root ganglion neurons
The Journal of physiology
579
:1–14
PubMed
Google Scholar
A single sodium channel mutation produces hyper-or hypoexcitability in different types of neurons
Proc Natl Acad Sci U S A
103
:8245–8250
PubMed
Google Scholar
A novel tetrodotoxin-sensitive, voltage-gated sodium channel expressed in rat and human dorsal root ganglia
J Biol Chem
272
:14805–14809
PubMed
Google Scholar
Quantitative differences in neuronal subpopulations between mouse and human dorsal root ganglia demonstrated with RNAscope in situ hybridization
Pain
161
:2410–2424
PubMed
Google Scholar
Lack of Detection of the Analgesic Properties of PF-05089771, a Selective Na(v) 1.7 Inhibitor, Using a Battery of Pain Models in Healthy Subjects
Clin Transl Sci
13
:318–324
PubMed
Google Scholar
Assessing NaV1.7 during tonic firing in pig C-nociceptors
PLoS One
20
:e0335081
PubMed
Google Scholar
Modulation of human dorsal root ganglion neuron firing by the Nav1.8 inhibitor suzetrigine
Proc Natl Acad Sci U S A
122
:e2503570122
PubMed
Google Scholar
Cutaneous pain in disorders affecting peripheral nerves
Neurosci Lett
765
:136233
PubMed
Google Scholar
Spatial transcriptomics of dorsal root ganglia identifies molecular signatures of human nociceptors
Sci Transl Med
14
:eabj8186
PubMed
Google Scholar
Interplay of Nav1.8 and Nav1.7 channels drives neuronal hyperexcitability in neuropathic pain
J Gen Physiol
156
PubMed
Google Scholar
Nav1.8: Intrinsic limits on the functional effect of abrogation in DRG neurons
Proc Natl Acad Sci U S A
122
:e2507342122
PubMed
Google Scholar
Nav1.8 and Chronic Pain: From Laboratory Animals to Clinical Patients
Biomolecules
15
PubMed
Google Scholar
Similar excitability through different sodium channels and implications for the analgesic efficacy of selective drugs
eLife
12
PubMed
Google Scholar
Discordance between preclinical and clinical testing of Na V 1.7-selective inhibitors for pain
Pain
166
:481–501
PubMed
Google Scholar
Single C nociceptor responses and psychophysical parameters of evoked pain: effect of rate of rise of heat stimuli in humans
J Physiol
450
:581–592
PubMed
Google Scholar
Voltage-gated Na(+) currents in human dorsal root ganglion neurons
eLife
6
:e23235
PubMed
Google Scholar
Article and author information
Author information
Cite all versions
You can cite all versions using the DOI
10.7554/eLife.111490
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2026,
Fujita et al.
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
views
0
downloads
0
citations
0
Views, downloads and citations are aggregated across all versions of this paper published by eLife.





