 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库实锤!耐力训练不仅练肌肉,还强大脑!Neuron最新研究:大脑中SF1神经元才是调控运动收益的核心,激活它们,运动效果翻倍

“练耐力不就是练腿、练心肺吗?”这几乎是所有人对运动的固有认知。我们总以为,跑不动是因为肌肉力量不够、肺活量不行,只要多跑、多练,身体自然会变强。但现实却常常打脸:同样坚持 3 个月跑步,有人能轻松完成半程马拉松,有人却连 3 公里都喘得不行;有人越练越精神,有人却越练越疲惫,陷入“无效运动”的怪圈。
长期以来,运动生理学也一直将研究重心放在肌肉、心血管、代谢等外周组织上,认为运动带来的所有收益,都是这些外周器官重塑的结果。大脑虽然会因运动产生神经发生、突触连接增加等变化,但这些都被视为运动的“副产品”,与耐力提升本身无关。
然而,最新发布在《Neuron》上的一项研究,彻底推翻了这一延续数十年的传统观点。来自宾夕法尼亚大学、德克萨斯大学西南医学中心等顶尖机构的科学家团队证实:耐力训练本质上是一场“大脑主导的全身重塑”,而大脑下丘脑腹内侧核(VMH)中一类表达类固醇生成因子 1(SF1)的神经元,才是调控运动收益的“总开关”。运动不仅能激活这些神经元,还会从结构和功能上对它们进行深度重塑;抑制它们的活动,再刻苦的训练也无法提升耐力;而精准激活它们,就能让运动效果直接翻倍!

研究背景与核心目的
过去半个世纪,科学家们清晰描绘出了运动如何改造外周身体:跑步能让肌肉纤维变粗、线粒体数量增加,让心脏泵血能力更强,让脂肪燃烧效率更高。人们甚至发现了多种由肌肉分泌的“肌因子”,认为正是这些外周信号介导了运动带来的全身健康收益。基于这一理论,全球药企一直在研发“运动模拟药”,希望通过模拟这些肌因子的作用,让无法运动的人也能获得运动的好处。
但越来越多的研究发现,这一理论存在无法解释的漏洞:同样的训练强度,不同人的收益天差地别,例如有些肌肉功能正常的人,却始终无法提升运动耐力,而敲除大脑中特定基因的小鼠,即使肌肉和心肺完全正常,也会表现出严重的运动能力缺陷。其中最关键的线索来自于 SF1 基因,此前研究发现,全身敲除 SF1 基因的小鼠,运动耐力会出现毁灭性的下降,但没人知道这一效应是否源于大脑中的 SF1 神经元。
由此,本研究的核心目的,就是解答这个悬而未决的问题:大脑中的 VMH SF1 神经元,是否是运动诱导耐力提升的必要且充分条件?运动如何通过改变这些神经元的特性,来调控全身的代谢和生理适应?
运动激活VMH SF1神经元
研究团队首先要验证的是:当我们运动时,大脑里有没有特定的神经元会被激活?他们把目光锁定在了下丘脑腹内侧核(VMH)中的 SF1 神经元上。这些神经元被称为身体的"能量管家",平时负责调控体重、血糖和能量消耗。
研究人员让小鼠进行一次力竭性跑步机运动,随后检测活动依赖性基因脑源性神经营养因子(Bdnf)的表达,这是神经元“兴奋过”后才会打开的标记基因。结果正如下图 1 所示:运动后的SF1神经元中,表达Bdnf的比例从34.5%上升到42.2%,而每个活跃神经元里的 Bdnf 转录本数量也增加了近一倍。这一结果清晰地证明:运动确实能特异性地唤醒大脑深处这群特殊的神经元。
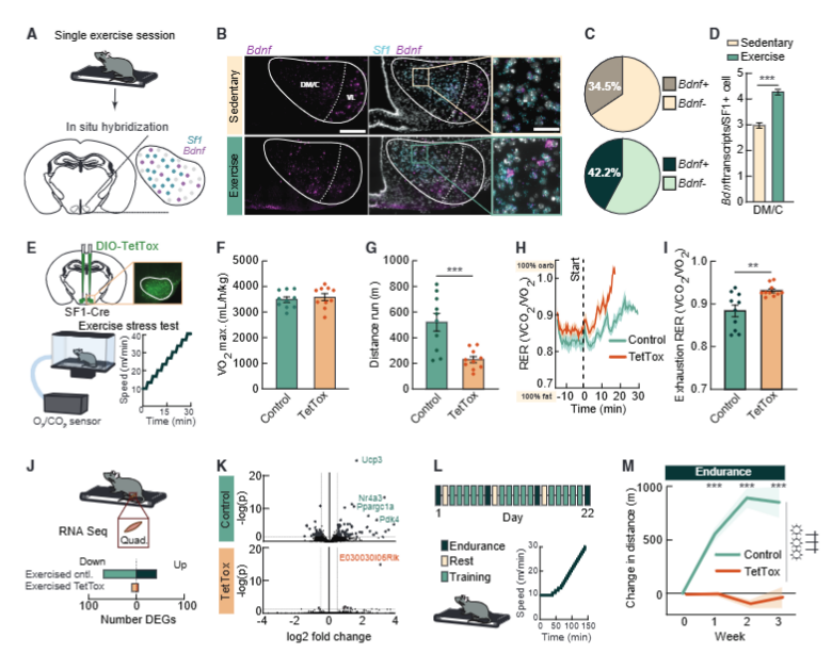
图1. VMH SF1输出是运动生理适应所必需的
切断“总司令”的通讯,再怎么练都白搭
既然运动能激活 SF1 神经元,那它们是不是真的在运动适应中扮演关键角色呢?为了找到答案,研究人员做了一个大胆的实验:他们利用基因编辑技术,在 SF1-Cre 小鼠的 VMH 脑区特异性表达破伤风毒素轻链(TetTox)。这种毒素不会损伤神经元本身,但会彻底切断它们和其他神经元的通讯,这相当于把“总司令”的通讯设备全关了,但身前线的士兵(肌肉、心肺)都完好无损。
结果非常明确:首先是运动表现的明显下滑。虽然 TetTox 小鼠的最大摄氧量和对照组完全相同,说明它们的心肺功能没有任何问题,但在耐力测试中,它们跑的距离更短、最快速度更低,更早出现疲劳。更让人意外的是它们的燃料利用模式出了问题:正常小鼠运动时,身体会循序渐进地从燃烧脂肪转向燃烧碳水化合物,以最大化能量利用效率,但 TetTox 小鼠在很低的运动强度下就匆忙切换成了烧糖模式,这也是它们更容易疲劳的重要原因。
最关键的发现来自于长期训练实验。经过 3 周规律的跑步机训练后,正常小鼠的耐力提升了近 1000 米,而 TetTox 小鼠的耐力几乎没有任何进步。对骨骼肌的转录组分析显示,正常小鼠运动后,上百个与线粒体生成、氧化代谢、肌肉重塑相关的基因会显著上调。但在 TetTox 小鼠中,这些训练诱导的基因变化几乎完全消失了。它们的肌肉仿佛“不知道”自己经历过训练,依然停留在久坐状态。
为了排除“初始运动能力差”和“体重差异”的干扰,研究人员还构建了 SF1 神经元特异性敲低 BDNF 的小鼠模型。这些小鼠天生运动能力更差、体重也更重,但仅仅训练 1 周后,它们的耐力就出现了显著提升。唯独 SF1 神经元通讯被切断的小鼠,无论怎么训练,都始终停留在原点,真正陷入了“练了白练”的困境。
运动让这群神经元越练越"兴奋"
为什么长期训练会让 SF1 神经元的作用越来越重要?
通过在体钙成像追踪,研究人员发现 SF1 神经元分为两个功能亚群:一类在运动中活跃、结束后抑制,另一类只在运动结束后激活。经过 1 周训练,跑后激活型神经元的比例从 31.8% 飙升至 53.2%,活动强度也明显增强,这说明训练会动态重塑神经元的响应模式,让更多神经元在运动后的关键适应期发挥作用。
进一步的电生理实验证实,运动后的 SF1 神经元兴奋性显著提升:静息膜电位更接近放电阈值,自发放电频率从 2.4Hz 增至 6.8Hz;久坐小鼠中 37% 几乎不活动的“沉默神经元”,在运动小鼠中全部被唤醒。
电生理的发现让研究人员好奇:为什么运动后的神经元会变得这么活跃?答案就藏在神经元的结构里!通过高分辨率显微镜观察,他们发现运动小鼠的 SF1 神经元树突上,树突棘的密度整整增加了一倍——树突棘就像是接收信号的小天线,数量越多,能接收的信号就越多。简单来说,运动就是给大脑的“运动指挥中心”扩建了更多信息通道,让整个指挥系统更灵敏、更高效。
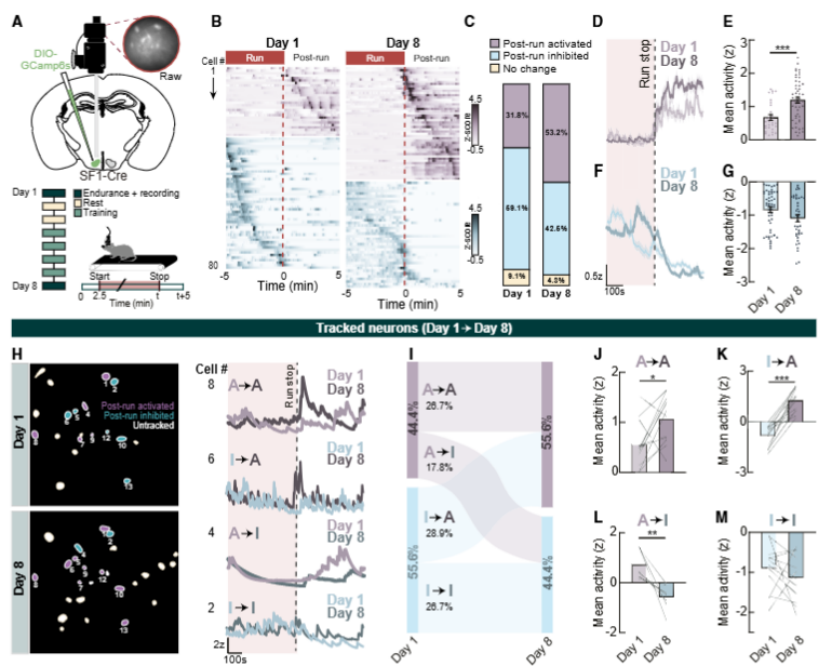
图2.运动训练增加SF1神经元激活
运动后"趁热打铁",刺激神经元让效果翻倍
基于这些发现,研究人员提出核心假设:运动后 SF1 神经元的持续激活,才是驱动训练收益的关键。他们通过光遗传技术做了双向验证:每次训练后暂时抑制 SF1 神经元 15 分钟,小鼠的耐力进步会被大幅削弱,相当于训练效果被强行 “暂停”;而每次训练后额外激活神经元 60 分钟,小鼠的耐力提升幅度远超普通组,甚至在普通组进入训练平台期后,激活组仍能继续突破上限。
值得注意的是,单独刺激这群神经元(不运动)就能提高血糖、增加碳水化合物利用和能量消耗,但这种刺激产生的身体活动量很小,意味着SF1神经元的价值不在于代替运动,而在于运动后的特定时间窗口内放大训练收益。
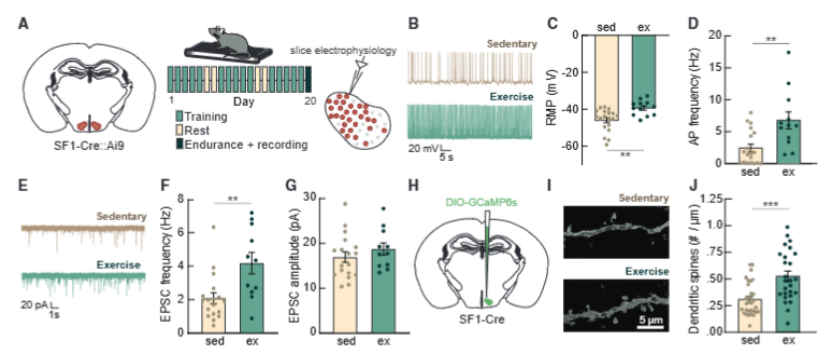
图3.运动训练增加VMH SF1神经元的兴奋性输入
原来,耐力训练从来不是单纯的“体力活”,而是一场大脑和身体的双向奔赴呢~
参考文献:
Kindel M, Post R, Grose K et al.Exercise-induced activation of ventromedial hypothalamic steroidogenic factor-1 neurons mediates improvements in endurance
Neuron, 2026; 114, 1564-1575.e9
撰文|ZZN
编辑 | lcc

点击下方「阅读原文」,前往生物谷官网查询更多生物相关资讯~





