 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库Starvation of the bacteriuminduces simultaneous attacks on the dinoflagellate
Abstract
Phytoplankton serve as a source of nutrients for bacteria in the marine environment. The interactions between algae and bacteria are known to include mutualism, commensalism, competition or antagonism. This occurs in the microenvironment surrounding phytoplankton cells, the phycosphere, an interface rich in nutrients and organic molecules exuded by the cell. Here, based on
in situ
observations and on an
in vitro
interaction study, we report on a novel form of starvation-induced hunting that the cells of selected Vibrio species exert on dinoflagellates. The results showed that
Vibrio atlanticus
was capable of attacking and killing the dinoflagellate
Alexandrium pacificum
ACT03. Briefly, the observed mechanism of algal-killing consists of first, the ‘immobilization stage’ involving the secretion of algicidal metabolites that disrupt the flagella of the algae. In the ‘attack stage’, Vibrios simultaneously surround algal cells at high density for a brief period without invading them. Finally, the ‘killing stage’ in which the lysis and consumption of the dinoflagellates occur. By using a combination of biochemical, proteomic, molecular and fluorescence microscopy approaches, we showed that this relationship is not related to the decomposition of algal organic matter, Vibrio quorum sensing pathways, toxicity of the algae or pathogenicity of the bacterium but is conditioned by nutrient stress, iron availability and linked to the iron-vibrioferrin transport system of
Vibrio atlanticus
. This is the first evidence of a new mechanism that could be involved in regulating Alexandrium spp. blooms and giving Vibrio a competitive advantage in obtaining nutrients from the environment. The interaction model we propose here suggests that Vibrio could play a role in regulating the proliferation of Alexandrium spp., giving it a competitive advantage in obtaining nutrients from the environment.
Introduction
Harmful algal blooms (HABs) have experienced an increase in their occurrence, intensity, and geographical distribution on a global scale, resulting in adverse environmental, health, and socioeconomic impacts (
Marampouti et al., 2021
). HABs have a considerable impact on human health as a result of direct exposure to volatile toxins or by toxic seafood consumption (
Burkholder et al., 2018
). From an ecological point of view, the expansion of HABs can result in the erosion of biodiversity, because they cause massive mortality of marine species and they are generally monospecific in nature (
Chai et al., 2020
). In coastal areas, understanding the biological interactions that control toxic algal blooms is therefore a major ecological challenge. Among HAB-causing organisms, a number of
Alexandrium
species have been placed on the list of invasive Mediterranean species. Among them,
Alexandrium pacificum
is a flagellated eukaryotic unicellular organism that together with
Alexandrium tamarense
and
Alexandrium fundyense
forms the "Alexandrium tamarense" complex responsible for paralytic shellfish poisoning worldwide (
Hadjadji et al., 2020
). Since 1998,
A. pacificum
(former
A. catenella
) was monitored by the French phytoplankton observation and monitoring network (Rephy) in the Thau lagoon (French Mediterranean) because it produces paralytic shellfish toxins (PSTs) resulting in paralytic shellfish poisoning (PSP) syndrome.
Laanaia (
Laanaia et al., 2013
) showed that in Thau lagoon, a water temperature around 20°C for several days and organic and inorganic nutrients in sufficient concentrations are parameters favoring the development of
A. pacificum
, whose massive blooms occur in autumn. Algal blooms are seasonal events resulting in a rapid increase in the concentration of a species of algae in an aquatic environment. Depending on the species of phytoplankton, tolerance to physicochemical parameters varies, which influences when blooms occur (
Leblad et al., 2020
). Interestingly, although the collapse of phytoplankton blooms has been previously attributed to viruses (
Pal et al., 2020
), some ecological studies have suggested an important role of algicidal bacteria (
Su et al., 2007
;
Wang et al., 2010
). Among them several are belonging to the Vibrio genus (
Li et al., 2014
;
Wang et al., 2020
).
Vibrio (class γ-proteobacteria) are common microorganisms in marine systems worldwide (
Baker-Austin et al., 2017
;
Mavian et al., 2020
), where they are important components of the food chain, particularly in biodegradation, nutrient regeneration and biogeochemical cycles (
Oberbeckmann et al., 2012
). Vibrio is one of the most studied bacterial taxa due to their ubiquity in coastal marine systems and their capacity to cause infections in humans and animals, leading sometimes to epizootic or zoonotic epidemics (
LeRoux et al., 2015
;
Mavian et al., 2020
). Vibrio are extremely adaptable to their environment (
Johnson, 2013
). The main factors influencing their occurrence and distribution in water are temperature, salinity, nutrient availability (
Wang et al., 2020
), multiple strategies such as biofilm formation on biotic and abiotic surfaces (
Espinoza-Vergara et al., 2020
), or interactions with a multitude of other organisms such as eukaryotic predators (
Drebes Dörr and Blokesch, 2020
) or plankton (
Lopez-Joven et al., 2018
) are used by Vibrio in the environment. There is also evidence that global climate change has increased Vibrio-associated illnesses affecting humans and animals (
Brumfield et al., 2021
;
Muhling et al., 2017
). However, the drivers and dynamics of Vibrio survival and propagation in the marine environment are not yet fully understood.
A substantial number of research articles have highlighted the potential of γ-proteobacteria to exert algicidal activity against dinoflagellates, supporting the hypothesis that γ-proteobacteria such as Vibrio play a role in the control of algal blooms
in situ
(
Coyne et al., 2022
). However, the mechanisms behind Vibrio-driven algal lysis in the environment remain to be elucidated. Particularly, it is unclear how in the water column, algicidal compounds secreted by bacteria can concentrate around the algae to exert their lytic effect.
This study aims to describe observations made in the natural environment between Vibrio bacteria and Alexandrium algal blooms, and to determine
in vitro
the main factors involved in this relationship. Using a combination of biochemical, proteomic, molecular, and fluorescence microscopy approaches, we explored the role in algal toxicity, bacterial pathogenicity and the quorum sensing pathway on this relationship and showed the important role of nutrient stress and the iron uptake pathway in this unique Vibrio/Alexandrium interaction.
Methods
Quantification of
Alexandrium
algae and Vibrio bacteria in the environment by qPCR
Seawater samples were collected in the Thau Lagoon (southern France, a shallow Mediterranean ecosystem open to the sea (
Abadie et al., 1999
), during spring and autumn 2015. Briefly, samples were collected from the subsurface (-50 cm) near an oyster table at a phytoplankton surveillance site (part of the REPHY network, N 43°26.058’ and E 003°39.878’). Once a week during spring and autumn 2015, during field sampling campaigns, 20 L of water was filtered on board through a 180 μm pore-size nylon membrane. At the laboratory, according to Lopez-Joven et al. (
Lopez-Joven et al., 2018
) seawater was fractionated into two size classes as follows: 2 L of the above filtrate was filtered through a 0.8 μm pore-size polycarbonate Whatman Nuclepore membrane to obtain organisms in the 0.8–180 μm range corresponding to plankton-associated Vibrio and living
Alexandrium
forms. Then, the filtrate from the 0.8 µm membrane was filtered again through a 0.2 μm pore-size polycarbonate Whatman Nuclepore membrane until the membrane was saturated.
Alexandrium
cells, ranging from 25 to 40 µm, belong to the microphytoplankton and are therefore retained in the 0.8–180 µm fraction. Any Vibrio cells potentially associated with or attached to
Alexandrium
cells will also be retained in this fraction. Vibrio cells are approximately 0.5-0.8 µm thick. The fraction between 0.2 and 0.8 µm therefore includes the free-living Vibrio. The bacterial population collected on 0.8-µm-pore-size filters was designated the particle-associated community, and the population on 0.2-µm-pore-size filters was designated as the free-living community. Membranes (in triplicate) were then conserved in 500 µL of 100% EtOH at -20°C. Environmental DNA (eDNA) was extracted from the MF Millipore membrane using the Macherey-Nagel NucleoSpin Tissue Kit and resuspended in 100 µL of water. The samples were then stored at -20°C after eDNA quantity and purity were assessed using a NanoDrop system (NanoDrop Technologies, Wilmington, DE, USA). PCR amplification reactions were done on a Roche LightCycler 480 Real-Time thermocycler (qPHD platform, University of Montpellier, France) using specific primer pairs (
Table 3
). Typically, the reactions contained 1 µL of template DNA (the DNA concentration for all samples varied from 1 to 40 μg mL
−1
), 0.5 µL of each primer (3.33 µM) and 4 µL of reaction mixture (SYBR Green Master Mix) in a total volume of 6 μL. The reaction parameters were as follows: 5 minutes at 95°C (initial denaturation) and 40 cycles of 10 s at 95°C (denaturation), 10 s at the corresponding hybridization temperature (
Table 3
) and 10 s at 72°C (elongation). Melting curve profiles were generated by increasing the temperature from 65°C to 95°C at 0.5°C s
-1
. Amplification products were analysed using LightCycler software (Roche Diagnostics).
Vibrio spp
. and
A. pacificum
and
A. tamarense
were quantified by constructing calibration curves based on DNA from the
V. atlanticus
LGP32 reference strain (former
V. tasmaniensis
LGP32) and from the
A. pacificum
reference strain (ACT03:
A. catenella
strain isolated from Thau in 2003) and the
A. tamarense
reference strain (ATT07:
A. tamarense
isolated from Thau in 2007) (not shown).
Strains and growth conditions
Vibrio strains. Wild-type and isogenic mutants of
V. atlanticus
LGP32 (
Table 1
) were used in this study. Deletion-mutants included Δ
luxR
, Δ
luxS
, Δ
luxM
and Δ
pvuB
isogenic strains. The Δ
pvuB
mutant was constructed here by allelic exchange as described previously by Le Roux (
Le Roux et al., 2007
). We also used
V. atlanticus
LGP32 carrying the pSW3654T-GFP plasmid (
Le Roux et al., 2007
), hereafter referred to as
V. atlanticus
LGP32-GFP. Bacterial strains were grown at 22 ± 1°C in Zobell medium (0.38 µM iron (III)). When needed, 25 μg mL
-1
chloramphenicol (Cm) was added to cultures of
V. atlanticus
LGP32-GFP (
Le Roux et al., 2007
).
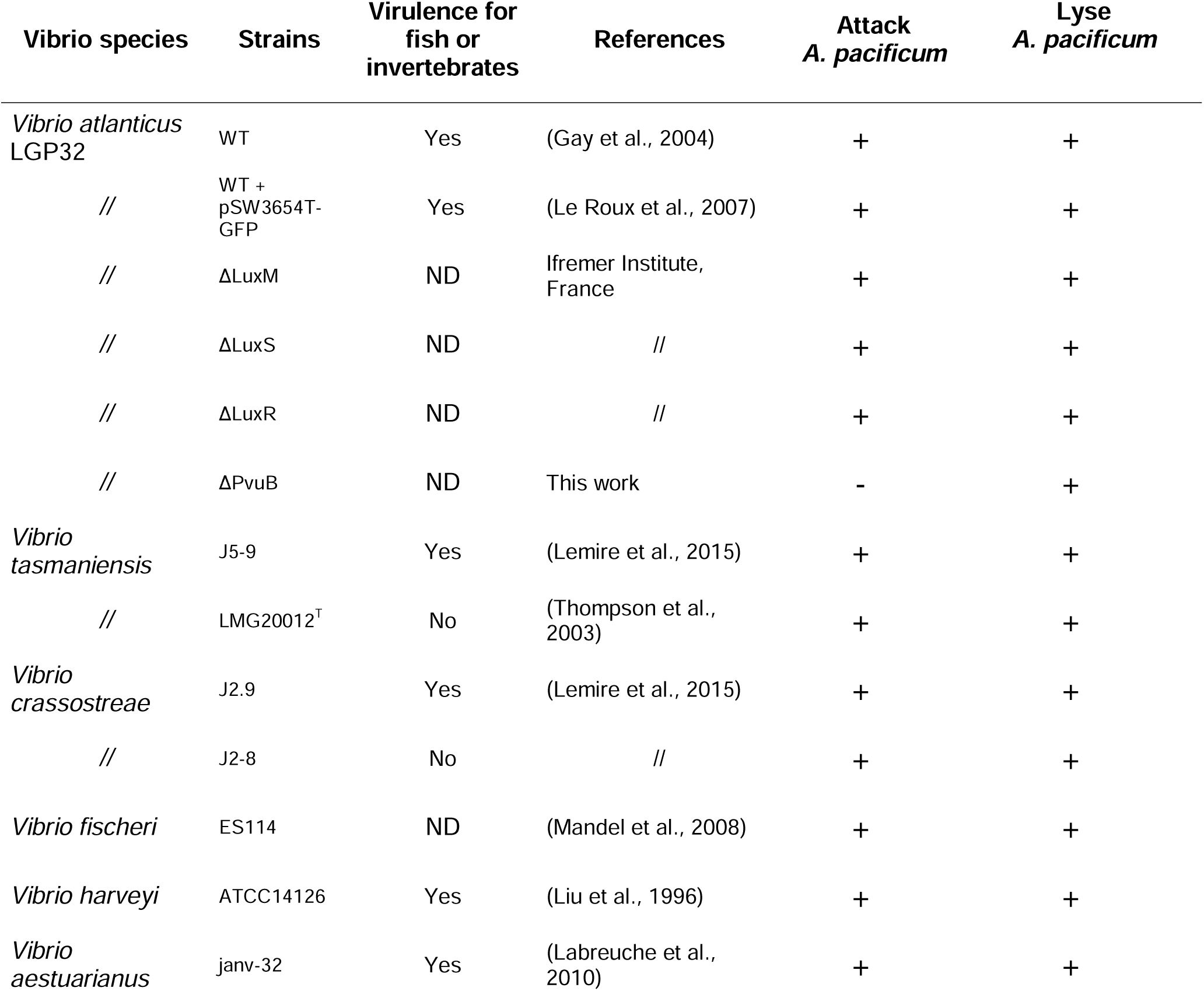
Ability of Vibrio strains to attack and to lyse
Alexandrium pacificum
.
ND: not determined.
Phytoplankton strains
. Non-axenic phytoplankton species (
Table 2
) were grown in batch culture in enriched natural sea water (ENSW, 6.55 µM iron (III)) with a salinity of 36 practical salinity units (PSU) at 22 ± 1°C under cool white fluorescent illumination (100 µmol photons m
-2
s
-1
) and a 12 h:12 h light:dark cycle (
Harrison et al., 1980
). The algae were used for experiments in their exponential growth phase.
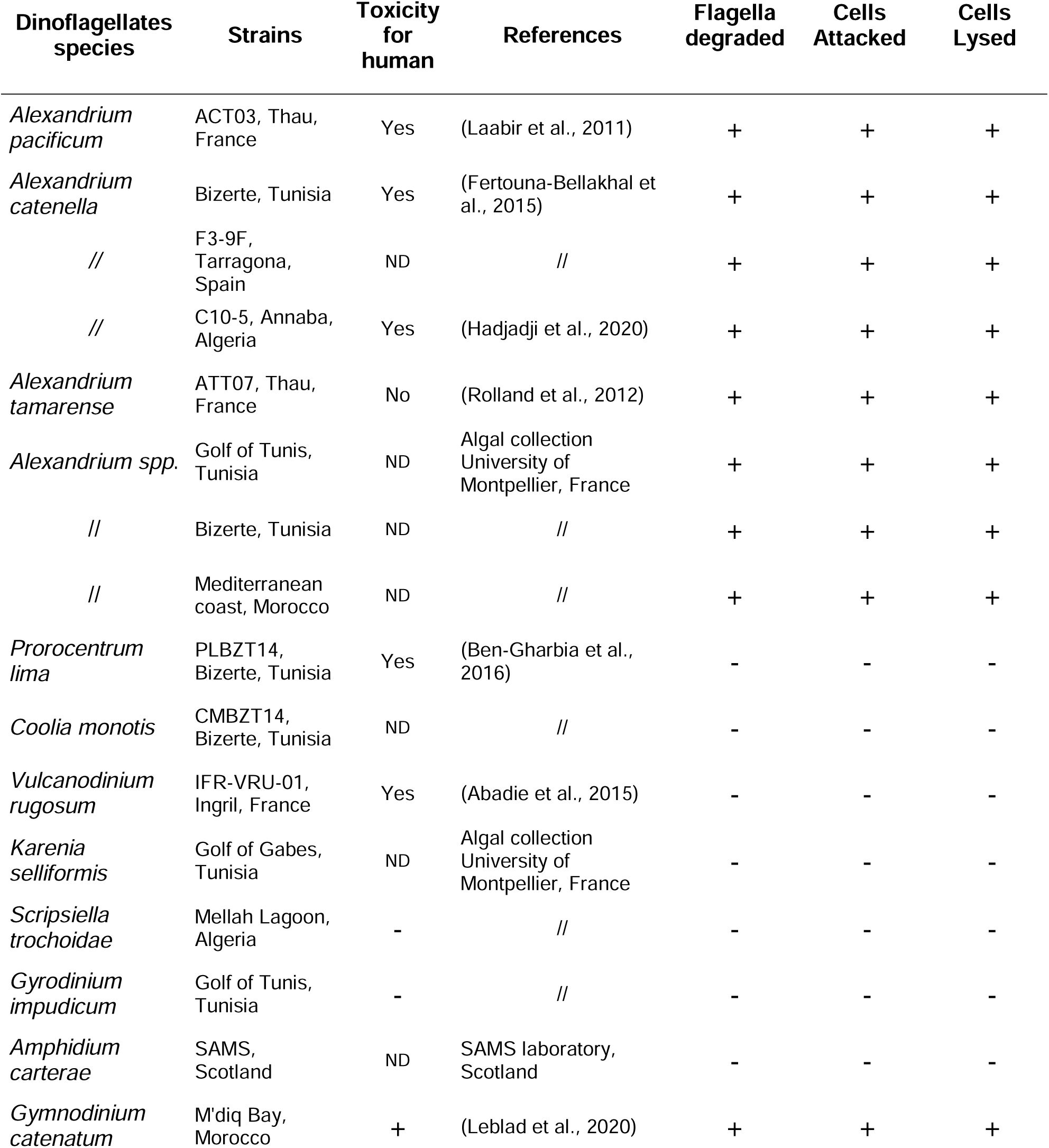
Ability of
V. atlanticus
LGP32 to degrade flagella, attack and lyse the targeted dinoflagellates
spp
. commonly found in the Mediterranean Sea.
ND not determined.
Co-culture assay
For each tested phytoplankton species (
Table 2
), 2 x 10
4
cells harvested in their exponential growth phase (doubling time between 5 and 7 days) were placed in 20 mL of ENSW medium in a 50 mL suspension culture flask (Cellstar® PS, Greiner bio-one). After incubation for 24 h at 22 ± 1°C under cool white fluorescent illumination, 40 µL of Vibrio strains (
Table 1
) grown for 12, 36, 60 and 156 h in Zobell medium or in Zobell medium supplemented with FeCl
3
(6 µM iron (III)) or with boron (H
3
BO
3;
0.47 mM) or the corresponding culture supernatant were added to the phytoplankton cells (
Table 2
). After incubation at 22 ± 1°C for 0, 15, 30, 45 and 60 min under cool white fluorescent lights, living, non-swimming, attacked and lysed phytoplankton cells were counted in a sedimentation chamber under an inverted microscope. The number of lysed cells corresponded to phytoplankton cells showing disrupted membranes. Non-swimming algae were not counted as lysed cells. For Vibrio analysis, 100 µL of a 1:10 serial dilution mixture in ENSW (from 10
-2
to 10
-10
) was plated on Vibrio Selective TCBS (thiosulfate-citrate-bile salts-sucrose) agar (in triplicate). After incubation for 24 h at 22 ± 1°C, the number of living Vibrio cells was determined by counting colony-forming units (CFUs). The data come from three independent experiments using independent phytoplankton cultures and independent bacterial cultures.
Microscope observations
The dynamic of the interaction between
A. pacificum
ACT03 isolated from the French Thau Lagoon, south of France (
Laabir et al., 2011
) and
V. atlanticus
LGP32, which is an oyster pathogen isolated from the French Atlantic coast (
Gay et al., 2004
) and present in the Thau Lagoon (
Lopez-Joven et al., 2018
) was surveyed. As the
A. pacificum
ACT03 strain (
Table 2
) used in the study is not axenic, there is potential for bacteria other than
V. atlanticus
LGP32 to be present in the experiments. To elucidate the interaction without thoroughly accounting for the non-axenic cells, interaction was observed under a Zeiss Axio upright fluorescence microscope equipped with an AxioCamMRm 2 digital microscope camera using
V. atlanticus
LGP32 tagged with green fluorescent protein (GFP). Lasers were used at excitation wavelength (λex) 488 nm for GFP (emission wavelengths (λem): 505–530 nm) and λex 532 nm for plankton chloroplasts (λem 560–630 nm). Images were taken sequentially to avoid cross-contamination between fluorochromes. Sequences of images were merged during the Vibrio-
Alexandrium
interaction using ZEN 2012 (blue edition) software. Interaction events between Vibrio strains and phytoplankton strains were also observed under a Leica TCS SPE confocal laser scanning system connected to a Leica DM 2500 upright microscope camera (Montpellier RIO Imaging Platform, University of Montpellier, France).
Comparative proteomic analysis
Vibrio sampling and protein extraction
V. atlanticus
LGP32 was grown for 60 h at 22°C in artificial seawater (high nutrient stress, cond. 1) or 12 h at 22°C in Zobell media (no nutrient stress, cond. 2). After 10 min centrifugation at 8000 rpm, crude protein extracts of
V. atlanticus
LGP32 in each culture condition (triplicates) were obtained by sonication on ice at 20% amplitude for 20 s in 200 µL of ice-cold denaturing buffer (7 M urea, 2 M thiourea, 4% CHAPS in 30 mM Tris-HCl, pH 8.5) and clarified by centrifugation at 2000 x
g
, 15 min, 4°C. The protein concentration of the supernatant was estimated using the 2D Quant Kit (Cytiva™, MERCK) and samples were stored at -80°C until use.
Two-dimensional gel electrophoresis (2D gel)
Protein extracts were individually analysed on 2D gel electrophoresis (6 gels per condition each corresponding to different biological replicates). To do so, 100 µg of proteins from each extract was added to rehydration buffer (7 M urea, 2 M thiourea, 4% CHAPS, 65 mM DTT) for a total volume of 350 µL. They were then individually loaded onto 17 cm isoelectric focusing strips (Bio-Rad) with a stabilized non-linear pH ranging from 3 to 10. Due to the high complexity of the protein profile in the acidic part (left) of the gel pH 3–10 (
Fig. S3
), we conducted additional ‘close-up’ analyses in gels using 17 cm isoelectric focusing strips (Bio-Rad) with a narrower, stabilized pH gradient ranging from 4 to 7. Strips were rehydrated passively for 5 h at 22°C, followed by active rehydration for 14 h under a 50 V current at 22°C (to help large proteins enter the strips). Thereafter, isoelectrofocusing was carried out using the following programme: 50 V for 1 h, 250 V for 1 h, 8000 V for 1 h and a final step at 8000 V for a total of 140 000 V.h with a slow ramping voltage (quadratic increasing voltage) at each step. Focused proteins were reduced by incubating the strip twice in equilibration buffer (1.5 M Tris, 6 M urea, 2% SDS, 30% glycerol; bromophenol blue, pH 8.8) containing DTT (130 mM) at 55°C. Then, they were alkylated by incubation with equilibration buffer containing iodoacetamide (135 mM) on a rocking agitator (400 rpm) at room temperature protected from light. Proteins were also separated according to their molecular weight (second dimension) on 12% acrylamide/0.32% piperazine diacrylamide gels run at 25 mA per gel for 30 min followed by 75 mA per gel for 8 h using a Protean II XL system (Bio-Rad). Gels were stained using an MS-compatible silver staining protocol and scanned using a ChemiDoc MP Imaging System (Bio-Rad) associated with Image Lab software version 4.0.1 (Bio-Rad).
Comparative bioinformatics analysis of 2D gels
Twelve gels (six per condition) were selected for comparative analysis on PD-Quest v. 7.4.0 (Bio-Rad) to identify changes in protein abundance between the proteomic profiles of
V. atlanticus
LGP32 cultured in contrasting nutrient conditions (ENSW/Zobell). Spots whose mean intensity across six replicates per strain was two times higher or lower than those from the other strain, with a
P
<0.01 (Mann-Whitney U-test), were considered significantly different in terms of abundance between the two conditions (quantitative difference). Differentially represented spots were then excised from the gels, destained, trypsin-digested and the obtained peptides were identified by tandem mass spectrometry (MS-MS) using the PISSARO platform facility (University of Rouen, France). To identify protein(s) present in each spot, the obtained peptides were compared with
V. atlanticus
LGP32 reference genome (
). The genes whose peptides matched strongly were retrieved and used for a BLASTx query against non-redundant databases to determine the protein identity of the best match. A gene was considered as strongly matched when at least two peptides matched the sequence with a coverage of > 6%. Their theoretical isoelectric point (pI) and molecular weights were also calculated using the Expasy server (
) to compare them with the location of the spot on the gel. Altogether, these complementary analyses made it possible to characterize the protein identity of each spot with confidence.
Gene expression analysis
Vibrio sampling and RNA extraction
V. atlanticus
LGP32 was grown in Zobell media for 36 h and 60 h at 22°C (decline phase of growth, nutrient stress) or for 12 h at 22°C (exponential growth phase, no nutrient stress). Total RNA was isolated from
V. atlanticus
LGP32 using the standard TRIzol method (Invitrogen Life Technologies SAS, Saint-Aubin, France) and then treated with DNase (Invitrogen) to eliminate genomic DNA contamination. After sodium acetate precipitation, the quantity and quality of the total RNA were determined using a NanoDrop spectrophotometer and agarose gel electrophoresis. Following heat denaturing (70°C for 5 minutes), reverse transcription was performed using 1 μg of RNA prepared with 50 ng μL
-1
oligo-(dT) 12–18 mer in a 20-μL reaction containing 1 mM dNTPs, 1 unit μL
-1
RNAseOUT and 200 units μL
-1
Moloney murine leukaemia virus reverse transcriptase (M-MLV RT) in reverse transcriptase buffer, according to the manufacturer’s instructions (Invitrogen Life Technologies SAS, Saint-Aubin, France).
PCR amplification
Amplification reactions were analysed using a Roche LightCycler 480 Real-Time thermocycler (Bio-Environnement platform, University of Perpignan, France). In this study, several PCR primer pairs were designed using Primer3 software (optimal primer size: 20 bases; Tm: 60°C; primer GC%: 50; 2GC clamp and product size range: 150–200 bp) and calibrated with
V. atlanticus
LGP32 genomic DNA (
Table 1
). To determine the qPCR efficiency of each primer pair used, standard curves were generated using seven serial dilutions of genomic DNA (10
-2
, 10
-3
, 10
-4
, 10
-5
, 10
-7
and 10
-8
) (not shown); the qPCR efficiencies of the tested genes varied between 1.85 and 2.08 (
Table 3
). For gene expression, reverse transcription was performed with 1 µg of total RNA using random hexamers and SuperScript IV reverse transcriptase (Invitrogen). The total qPCR reaction volume was 10 μL and consisted of 5 μL of cDNA (diluted 1:5), 2.5 μL of SensiFAST SYBR No-ROX Mix (Bioline) and 100 nM or 300 nM PCR primer pair (
Table 3
). The reaction parameters were as follows: 2 min at 95°C (initial denaturation) and 40 cycles of 5 s at 95°C (denaturation), 10 s at 59°C (annealing) and 20 s at 72°C (elongation). The specificity of each PCR was checked by measuring fluorescent signals during melting curve analysis (PCR product heated from 65°C to 95°C continuously and slowly at 0.1°C s
−1
). Relative expression was calculated by normalization to the expression of two constitutively expressed housekeeping genes, namely, 6PKF (VS_2913) and CcmC (VS_0852), using the delta-delta threshold cycle (ΔΔCt) method (
Pfaffl, 2001
).
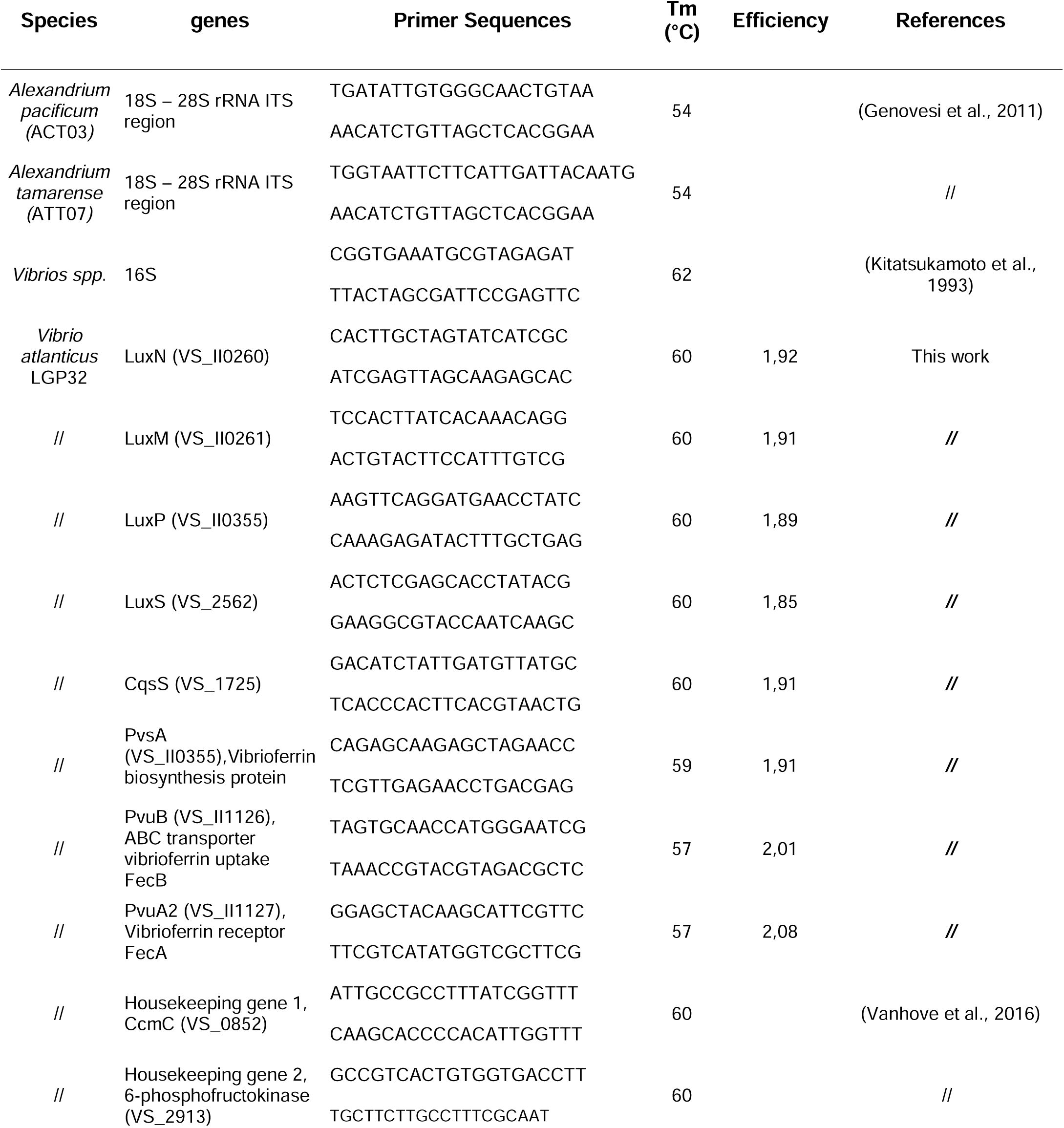
Oligonucleotide sequences of primers used for RNA expression analysis.
Detection of quorum-sensing signalling molecules
Vibrio culture
To detect the QS molecules (AI-2, AI-1 and CAI-1)
V. atlanticus
LGP32 was grown in Zobell media for 12 h (exponential growth phase, control) and 60 h (decline phase of growth, nutrient stress).
AI-2 analysis
Bioluminescence assay using the QS bioluminescent of
Vibrio campbellii
MM32 (
luxN
::Cm,
luxS
::Tn5Kan) was used to detect AI-2 molecules in culture supernatants. Briefly,
Vibrio
cultures were centrifuged at 17,000 x
g
for 10 min, and the resulting supernatants were filtered on 0.22 μm. Then 20 μL of the filtrates were mixed with 180 μL of
V. campbellii
MM32 diluted 1:5000 then incubated at 30°C and 100 rpm. Luminescence and cell density (OD620) were collected in triplicate and analysed according to Tourneroche et al.(
Tourneroche et al., 2019
).
AI-1 and CAI-1 extraction and LC-MS analysis
Chemical analyses were conducted with a Q Exactive Focus Orbitrap System coupled to an Ultimate 3000™ ultrahigh-performance liquid chromatography (UHPLC) system (Thermo Fisher Scientific) according to Rodrigues et al. (
Rodrigues et al., 2022
). Briefly, ethyl acetate (2 mL) was added into each culture (2 mL). This mixture was shaken overnight at room temperature (150 rpm). The two phases were then separated and the aqueous phase was extracted once again. The two obtained organic phases were pooled and the solvent was evaporated under vacuum. The crude extracts were dissolved in 500 µL LC-MS grade methanol for analysis. The experiments were performed using biological triplicates, each of which was analyzed in triplicate. Analyses of extracts and standards (3 μL injected) were performed in electrospray positive ionization mode in the 50–750 m/z range in centroid mode. The parameters were as follows: spray voltage: 3 kV; sheath flow rate: 75; aux gas pressure: 20; capillary temperature: 350°C; heater temperature: 430°C. The analysis was conducted in Full MS data-dependent MS2 mode (Discovery mode). Resolution was set to 70,000 in Full MS mode, and the AGC (automatic gain control) target was set to 1x10
6
. In MS2, resolution was 17,500, AGC target was set to 2x10
5
, isolation window was 0.4 m/z, and normalized collision energy was stepped to 15, 30 and 40 eV. The UHPLC column was a Phenomenex Luna Omega Polar C18 1.6 μm, 150 x 2.1 mm. The column temperature was set to 42°C, and the flow rate was 0.4 mL min
-1
. The solvent system was a mixture of water (A) with increasing proportions of acetonitrile (B), with both solvents modified with 0.1% formic acid. The gradient was as follows: 1% B 3 min before injection, then from 1 to 15 min, a gradient increase of B up to 100% (curve 5), followed by 100% B for 5 min. The flow was injected into the mass spectrometer starting immediately after injection. All data were acquired and processed using FreeStyle 1.5 software (Thermo Fisher Scientific).
Chemicals and solvents
N-acyl-homoserine lactones (AHL) were obtained from Cayman Chemical (Ann Arbor, MI, USA). Stock solutions were obtained by dissolving standards in methanol or dichloromethane (C18-AHL) at a concentration of 1 mg mL
-1
and stored at -80°C. Standard solutions for UHPLC- high-resolution tandem mass spectrometry (HRMS) analyses were prepared by diluting each individual standard solution with methanol in order to obtain a concentration range from 2000 to 20 ng mL
-1
. LC-MS grade methanol, acetonitrile and formic acid were purchased from Biosolve (Biosolve Chimie, Dieuze, France), analytical-grade ethyl acetate was obtained from Sigma-Aldrich. Pure water was obtained from Elga Purelab Flex System (Veolia LabWater STI, Antony, France).
Nature of lytic compounds secreted by
V. atlanticus
LGP32
To determine the temperature-sensitivity of the lytic compounds secreted,
V. atlanticus
LGP32 grown for 60 h in Zobell media at 22 °C was filtered through a 10 kDa membrane (Amicon
®
Ultra-4 filter unit). The eluate containing molecules with MW below 10 kDa was then incubated in a water bath at 100°C for 30 min. Boiled filtrates (0.1 mL) were subsequently used to inoculate
A. pacificum
(ACT03 strain) cultures, then lytic activity was observed under the Leica TCS SPE confocal laser scanning system. Zobell media with the same treatment was used as control.
Statistical analysis
Environmental data
Statistical analyses were performed using R 3.6.3 software (
R Core Team, 2024
) using Rstudio. The relationship between
Alexandrium
and Vibrio was explored separately in spring and autumn. We used a generalized linear model specifying a Gaussian family. For spring, the dataset for salinity and temperature was complete (14 periods of observation). An influential period was detected and removed from the dataset because no dead
Alexandrium
cells were observed. The effects of explanatory variables such as log10 (Vibrio+1), salinity and temperature were centred, reduced and tested as fixed effects with a linear relationship. Model selection was performed using the Akaike information criterion corrected for small sample size (AIC
c
). Models were considered different whenever the difference between their AIC
c
value and the lowest AIC
c
value (ΔAIC
c
) was lower than 2 (
Burnham and Anderson, 2002
).
Alexandrium
distribution and model residuals were checked for normal distribution assumptions (QQ plot and Shapiro-Wilk test). For autumn, the dataset was complete for 10 periods of observation. Salinity and temperature were missing for three periods. We explored the relationship between
Alexandrium
and Vibrio alone using the method detailed above for spring.
In vitro data
Statistical analyses were performed using one-way ANOVA (analysed by pair) followed by Tukey’s test (Statistica 10.0 software, StatSoft, Maison-Alfort, France). *P <0.05, **P <0.01, ***P <0.001.
Results
Concomitant occurrence of
A. pacificum
ACT03
, A. tamarense
ATT07 and free-living
Vibrio spp.
in the Thau lagoon
In the spring and autumn of 2015, in the Thau Lagoon (
Fig. 1A
), we detected
Alexandrium
algae (
A. pacificum
ACT03
and A. tamarense
ATT07, both alive and in degraded cell forms) and free-living Vibrio, but no plankton-associated Vibrio were observed (
Fig. 1B
, Data S1). Using model selection based on AICc, we found no significant relationship between
Alexandrium
(
A. pacificum
ACT03
, A. tamarense
) and
Vibrio spp.
abundances in autumn. This result is consistent with Vibrio’s difficulty in growing at temperatures below 20°C, as well as with the many environmental factors that can influence the dynamics of algae proliferation (
Laanaia et al., 2013
). Interestingly, in spring 2015, the mean densities of all
Alexandrium
cells (degraded and alive) and of free-living Vibrio were positively correlated. The lowest AICc was obtained with the model explaining degraded form of
Alexandrium
density based on the free Vibrio density (
Fig. 1C
). Given that, this model is not so different from the model with only the intercept, but better than any other linear combination with other potentially interfering drivers, such as temperature and salinity (
Fig. 1D
), we searched for evidence of a relationship between Vibrio and
Alexandrium
by studying their interaction
in vitro
.
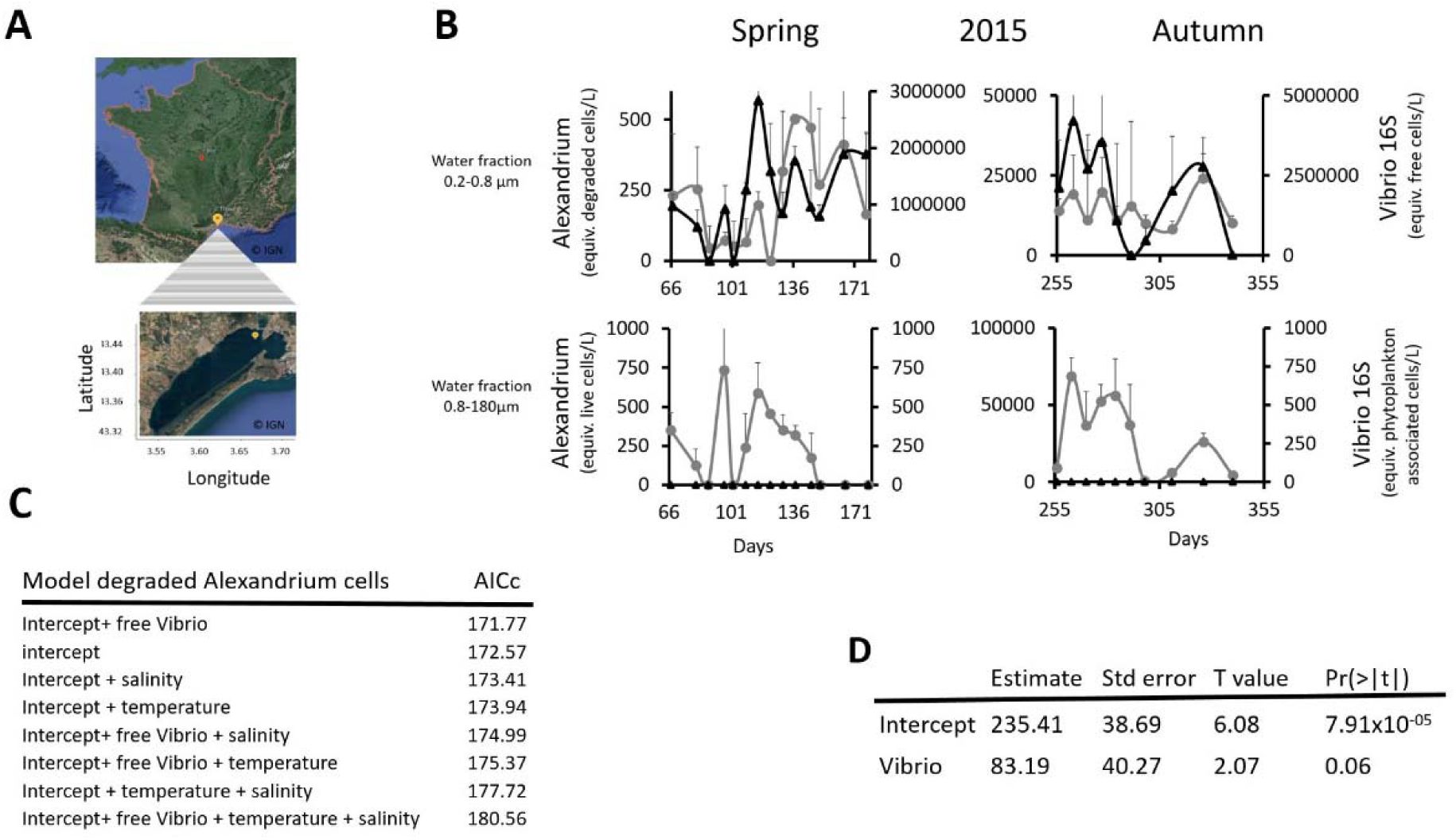
Dynamics of
Alexandrium
and Vibrio in the environment.
(A)
Location of the monitoring station in the Thau Lagoon (southern France).
(B)
Mean abundance (DNA equiv.) of
Vibrio spp
. (16S) and Alexandrium
spp
.
(A. pacificum
ACT03
+ A. tamarense
ATT07). Vibrio cells (black line with diamond dot) and degraded
Alexandrium
cells (grey line with round dot) were detected in the 0.2–0.8 μm fraction (free Vibrio fraction) in spring and autumn 2015. Living
Alexandrium
cells (grey line with round dot) but no plankton-associated
Vibrio spp
. (black line with diamond dot) were evidence in the 0.8–180 μm in spring and autumn.
(C)
Results of the Akaike Information Criterion (AICc) test conducted to select a model for explaining the mean value of dead Alexandrium (degraded cells) in spring.
(D)
Wald test of the AICc model explaining the mean value of dead Alexandrium in spring by free Vibrio.
V. atlanticus LGP32 feeds on Alexandrium pacificum ACT03
To investigate whether
Alexandrium
interacts with Vibrio, we incubated in Enriched Natural SeaWater (ENSW)
A. pacificum
ACT03 (2.0 x 10
4
cells) with
V. atlanticus
LGP32 previously grown for 12 hours in Zobell media (initial concentration of 8.8 x 10
7
cells mL
-1
). In interaction with
V. atlanticus
LGP32,
A. pacificum
ACT03 cell abundance decreased significantly from 2.1 x 10
4
cells mL
-1
after 1h of exposure to 1.1 x 10
4
cells mL
-1
after 48 h of exposure (
Fig. 2A
), while the
V. atlanticus
LGP32 concentration grew significantly after 26 h of interaction, reaching a maximum peak density of 7.6 x 10
7
CFU mL
-1
at 34 h (
Fig. 2B
). In the control experiment where
A. pacificum
ACT03 was cultured alone in ENSW, the algal concentration remained stable over time (
Fig. 2A
) and no bacteria were on the corresponding TCBS plates (Vibrio selective medium). In the control where
V. atlanticus
LGP32 was grown alone in ENSW, the bacterial concentration decreased from 7.0 × 10L CFU/mL after 1 hour of incubation to 1.1 × 10L CFU/mL after 48 hours of incubation (
Fig. 2B
). These results show that the interaction between
V. atlanticus
LGP32 and
A. pacificum
ACT03 leads to a decline in the algal population and promotes the growth of
V. atlanticus
LGP32. This suggests that
V. atlanticus
LGP32 is able to feed on
A. pacificum
ACT03.
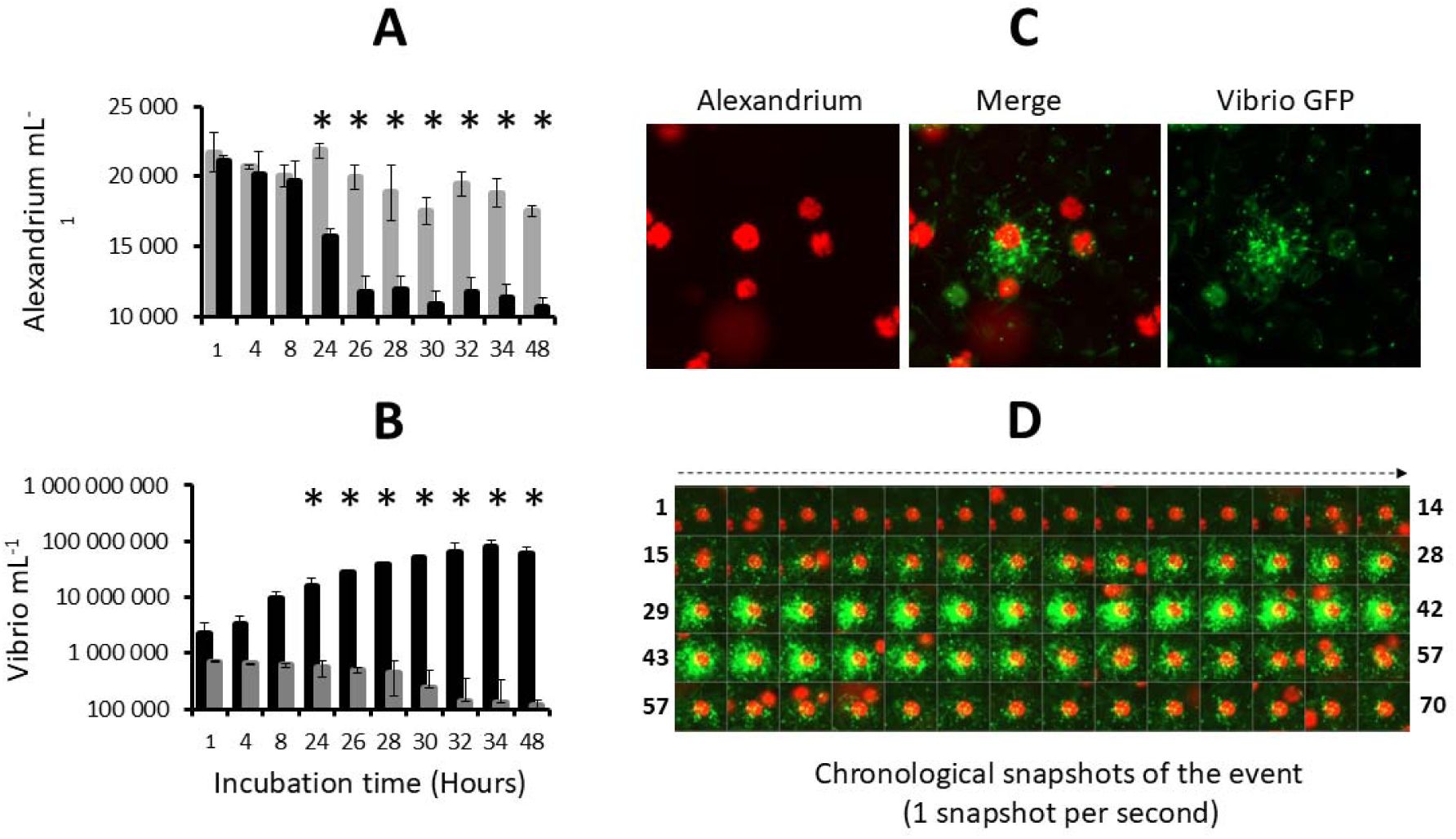
Incubation of
V. atlanticus
LGP32 and
Alexandrium pacificum
ACT03 in enriched natural seawater (ENSW).
(A)
A. pacificum
ACT03 cultured alone (grey bar) and incubated with
V. atlanticus
LGP32 (black bar) in ENSW.
(B)
V. atlanticus
LGP32 cultured alone (grey bar) and incubated with
A. pacificum
ACT03 (black bar) in ENSW.
(C)
Snapshot of the interaction between
V. atlanticus
LGP32-GFP cells (60-hour culture) and one cell of
A. pacificum
ACT03 taken at 8h00 of co-culture.
(D)
Chronological snapshots of the interaction (70 pictures, one per second).
V. atlanticus
LGP32 (small green cells) and
A. pacificum
ACT03 cell (large red cell). All experiments were done in triplicate. Asterisks indicate significant differences in a multiple comparison test (One-way ANOVA with post hoc Tukey test), *P ≤ 0.05.
V. atlanticus
LGP32 performs attacks on
A. pacificum
ACT03
Epifluorescence microscopy observation of GFP-labelled
V. atlanticus
LGP32 (previously grown in Zobell medium) in interaction showed that
V. atlanticus
LGP32 cells are capable of simultaneously attacking
A. pacificum
ACT03 cells (
Fig. 2C
and
Video 1
). The attacks were extremely rapid, with empty thecae (algal envelopes) observed in the medium after less than 60 s (
Fig. 2D
and
Video 2
). During the attack,
V. atlanticus
LGP32 did not invade the algal cell but remained clustered on the cell surface (
Fig. 2C
).
Attack of
A. pacificum
ACT03 is activated by
V. atlanticus
LGP32 starvation
In 2002, Martin hypothesized that nutritional stress induces bacteria to lyse algae (
Martin, 2002
). To test this hypothesis, we monitored
V. atlanticus
LGP32 behaviour in response to starvation (
Fig. 3
). We observed that
V. atlanticus
LGP32 in exponential growth phase (12 h of culture in Zobell medium, 8.8 x 10
7
cells mL
-1
) did not interact with
A. pacificum
ACT03 cells for the first hour of contact (
Fig. 3A, B, C
). In contrast,
V. atlanticus
LGP32 in the decline phase (36 h of culture in Zobell medium, 1.3 x 10
7
cells mL
-1
) induced a significant decrease in the number of motile algae cells by 8.9% after 15 min and by 43.3% after 60 min (
Fig. 3A
). This phenomenon corresponded to the degradation and/or disruption of algal flagella (
Video 3
). The flagella no longer functioned correctly, which caused irregular swimming of the algae (
Video 3
, left cell). This was followed by a complete cessation of swimming. When the flagellum detached from the algae (
Video 3
, right cell), the attack occurred. With starved
V. atlanticus
LGP32 (60 h of culture in Zobell medium, 0.6 x 10
7
cells mL
-1
), algae immobilization was fast and significant (91.4% in 15 min,
Fig. 3A
), and algae were attacked individually being targeted by
V. atlanticus
LGP32 cells (
Video 4
). The percentage of cells attacked and killed peaked at 30% after 15–30 min of contact (
Fig. 3B
and 3B1) and then decreased. After 1 h, attacks had stopped with approximately 40% of the algal cells still alive (
Fig. 3B1
and
3C
). Although it remains unclear whether the attacks occur during a specific phase of growth, it is evident that the cells are already weakened before attack as they have all lost their flagella. An old-starved culture of
V. atlanticus
LGP32 (126 h of culture in Zobell medium, < 0.1 x 10
7
cells mL
-1
) significantly immobilized
A. pacificum
ACT03 cells within a few minutes, with lysis occurring immediately (
Fig. 3A
and
3C
), making it impossible to detect attacks by
V. atlanticus
LGP32 (
Fig. 3B
). The lysis phase corresponded to initial vesicle formation followed by the bursting of
A. pacificum
ACT03 cells (
Fig. 3C
and
3C1
). Importantly, Vibrio densities decreased with culture age, ruling out the possibility that the stronger predation observed in older cultures was driven by higher bacterial densities.
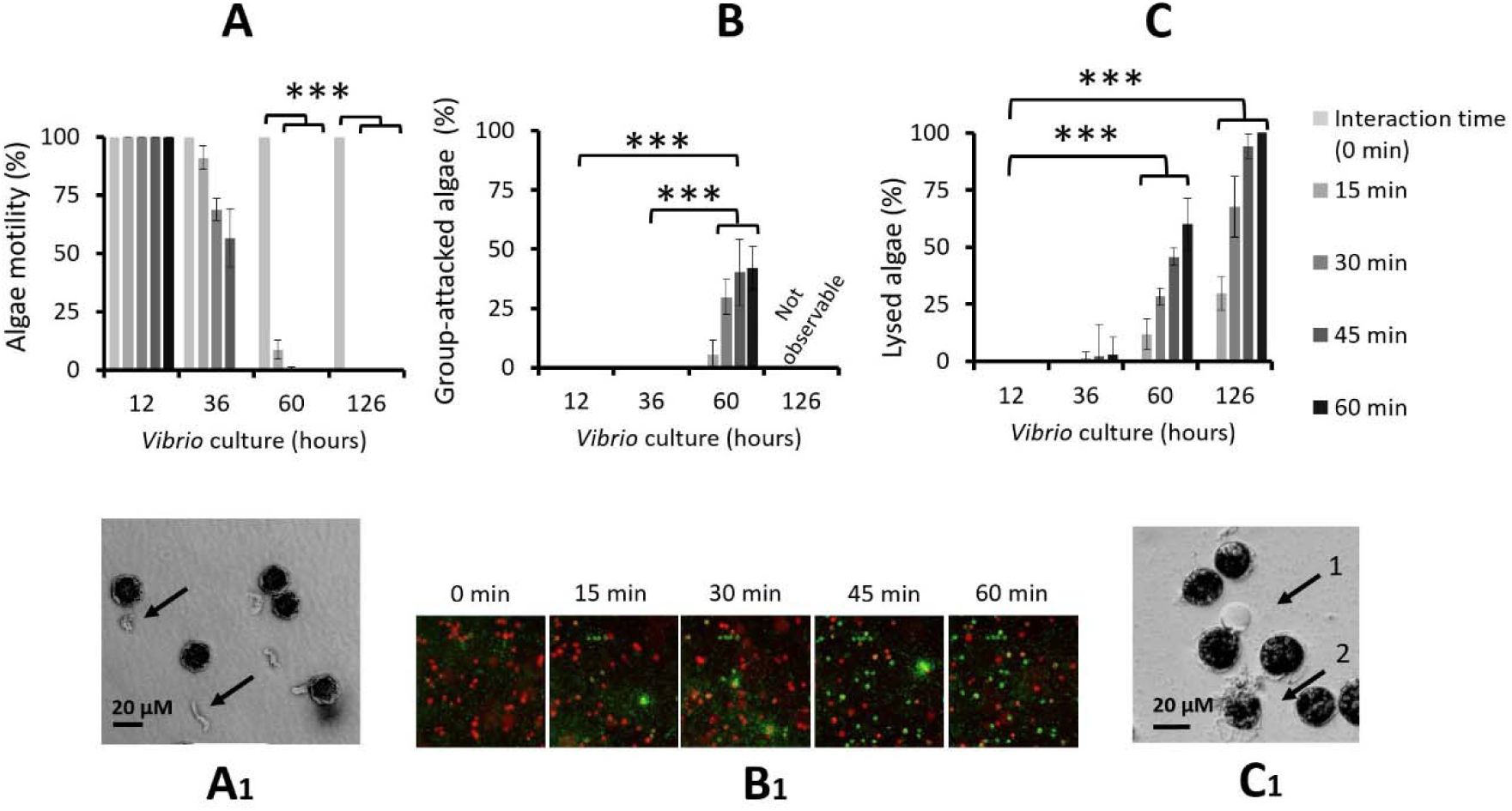
Role of
V. atlanticus
LGP32 starvation in the interspecific interaction process.
Experiments were conducted by incubating
A. pacificum
ACT03 with
V. atlanticus
LGP32 previously grown for 12, 36, 60 and 126 h in Zobell medium.
(A)
Cumulative percentage of motile
A. pacificum
ACT03 cells.
(B)
Cumulative number of cells attacked by
V. atlanticus
LGP32 and
(C
) Cumulative cell lysis after 0, 15, 30, 45 and 60 minutes of interaction. Corresponding pictures showing
(A1)
Black arrows indicate unhooked and degraded flagellum from
A. pacificum
ACT03 flagellum,
(B1)
Chronological sequence of five snapshots showing
V. atlanticus
LGP32-GFP cells (60-hour culture) and
A. pacificum
ACT03 cells, during the first hour of their interaction.
V. atlanticus
LGP32 (small green cells), living
A. pacificum
ACT03 (large red cells) and dead
A. pacificum
ACT03 (large green cell).
(C1)
Black arrow 1 indicates vesicle formation on
A. pacificum
ACT03 cell and black arrow 2 indicates lysed
A. pacificum
ACT03 cell. All percentages were determined based on a minimum of 2,000 cells of
A. pacificum
ACT03. All experiments were done in triplicate. Asterisks indicate significant differences in a multiple comparison test (One-way ANOVA with post hoc Tukey test), ***P ≤ 0.001
We next tested whether this lytic effect was linked to Vibrio culture supernatant and mediated by thermostable molecule(s) secreted by Vibrio. The culture supernatant of starved culture of
V. atlanticus
LGP32 (36 h) filtered through a 10 kDa membrane and then incubated at 100°C for 30 min still possessed its lytic properties, indicating that the algicidal compounds produced by
V. atlanticus
LGP32 are small thermostable molecules unlikely to be lytic enzymes, or lysins able to digest the algae cell. However, these experimental observations clearly show the key role of nutrient limitation in triggering the attack behaviour and the secretion of lytic compounds of
V. atlanticus
LGP32.
Attack occurs on
A. pacificum
ACT03 in exponential phase of growth
Here, we wondered whether the live/dead status of algae is important for
V. atlanticus
LGP32-mediated attacks targeting. To this end,
A. pacificum
ACT03 in exponential growth phase was first exposed for 30 minutes to the supernatant of a 126-hour culture of
V. atlanticus
LGP32, which induced lysis of 70% of the
A. pacificum
ACT03 cells (
Figures 3C
and
3C1
(arrow 2) and
Video 4
). Next, cells of
V. atlanticus
LGP32 from a 60-hour culture, capable of attacking
A. pacificum
ACT03 cells (
Fig. 3B
), were added. For 1 hour of exposure, no attack was observed on the previously lysed algae. This result is similar to what is observed on the
Video 1
with flash attacks only on immobilized, but not degraded
A. pacificum
ACT03 cells (red), and not on lysed cells (green). In addition, no attacks occurred on cells from an old
A. pacificum
ACT03 culture (1-month culture). Together, with the very short duration of attacks (
Video 1
), these results indicate that
V. atlanticus
LGP32 attacked exponentially growing cells of
A. pacificum
ACT03, but not decomposing algae, suggesting that this behaviour is not just an opportunistic response of heterotrophic bacteria to organic substrates.
Attack is independent of quorum sensing
Considering the simultaneous action of
V. atlanticus
LGP32 in attacks on
A. pacificum
ACT03, we tested whether the attack process depended on the key physiological mechanism that regulates many functions in marine microbial cells, quorum sensing (QS) (
Lami, 2019
;
Papenfort and Bassler, 2016
), a type of cell-cell communication. Although QS is a cell-density-dependent mechanism, our results showed no attack from a 12 h culture of
V. atlanticus
LGP32 up to a concentration of 4 x 10
6
Vibrio mL
-1
(
Fig 4A
). Attacks were only observed with
V. atlanticus
LGP32 from a 60 h culture at low concentration of 5 x 10
3
Vibrio mL
-1
to the highest concentration tested of 5 x 10
5
Vibrio mL
-1
(
Fig 4A
), consistent with the hypothesis that the attacks were independent of Vibrio density.
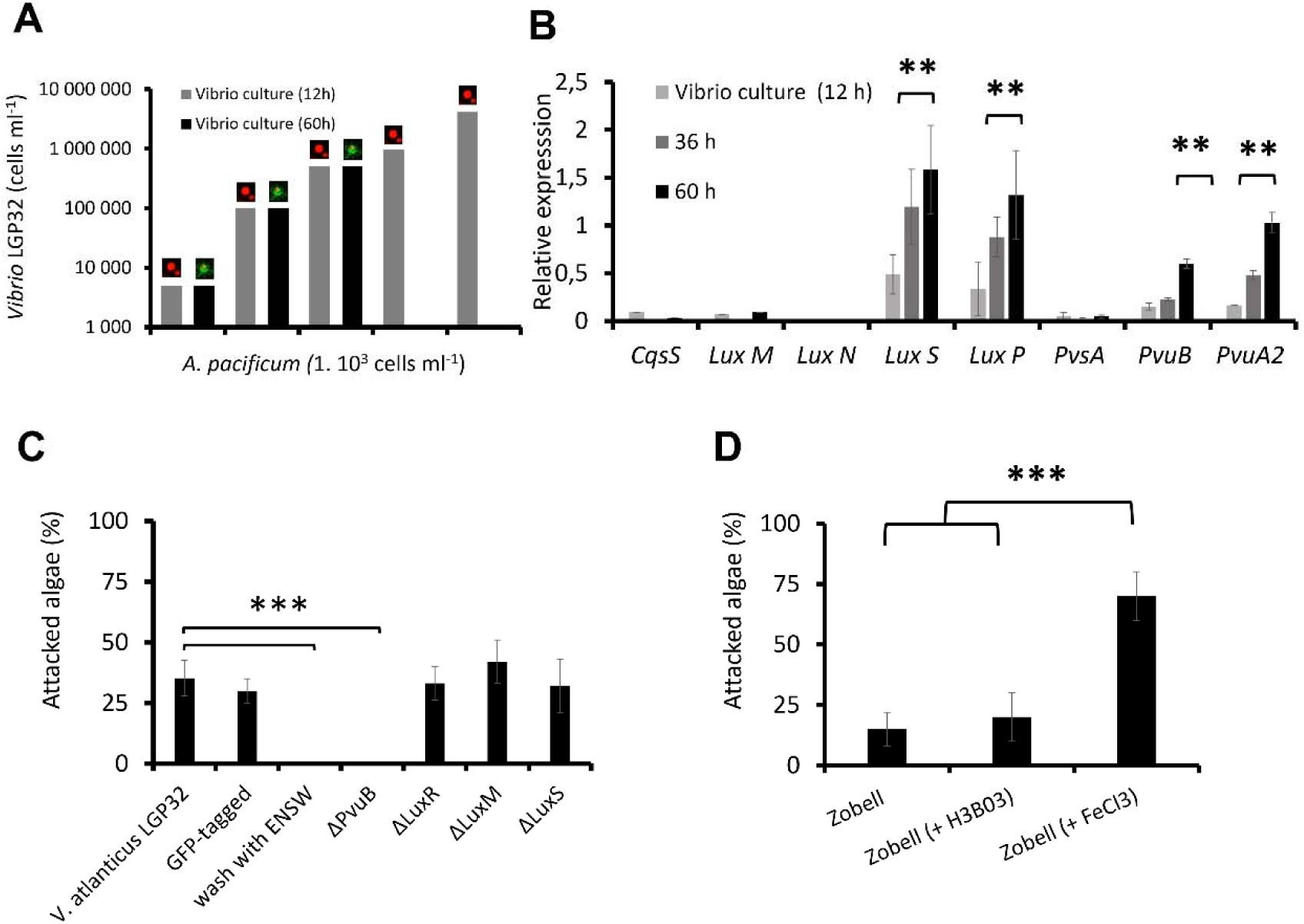
Role of quorum sensing and the vibrioferrin iron uptake pathway in the interaction process.
(A)
Effect of
V. atlanticus
LGP32 cell density on the attack process.
A. pacificum
ACT03 cells (1 x 10
3
cells mL
-1
) were incubated with
V. atlanticus
LGP32 grown for 60 hours in Zobell medium at concentrations ranging from 5.10
3
to 5 x 10
5
cells mL
−1
(black bars). For comparison,
A. pacificum
ACT03 incubated with
V. atlanticus
LGP32 grown for 12 hours in Zobell medium at concentrations ranging from 5 x 10
3
to 4 x 10
6
cells mL−1 (grey bars). The image on the bars indicates either unaffected algae (live red algae) or algae attacked by Vibrio (algae covered with green vibrio cell) during the interaction
(B)
CqsS, luxM, luxN, luxS, and luxP quorum sensing and PvsA, PvuB and PvuA2 vibrioferrin pathway genes expression in
V. atlanticus
LGP32 grown for 12, 36 and 60 h in Zobell medium.
(C)
Effect of
V. atlanticus
LGP32 mutants on the attack process. Experiments were conducted by incubating
A. pacificum
ACT03 with
V. atlanticus
LGP32,
V. atlanticus
LGP32 tagged with GFP,
V. atlanticus
LGP32 washed with ENSW or
V. atlanticus
LGP32 mutant ΔPvuB, ΔluxM, ΔluxR and ΔluxS previously grown 60 h in Zobell media (control), The percentage of
A. pacificum
ACT03 attacked was determined during the first 30 min of exposure.
(D)
Effect of
V. atlanticus
LGP32 culture media composition on the attack process. Experiments were conducted by incubating
A. pacificum
ACT03 with
V. atlanticus
LGP32 grown 60 h in Zobell media supplemented with boron (H
3
BO
3
) or FeCl
3.
The results were compared with an exposure to
V. atlanticus
LGP32 grown 60 h in Zobell media. All percentages were determined based on a minimum of 2,000 cells of
A. pacificum
ACT03. All experiments were done in triplicate. Asterisks indicate significant differences in a multiple comparison test (One-way ANOVA with post hoc Tukey test), **P ≤ 0.01, ***P ≤ 0.001.
The analysis of the expression of genes involved in the known QS pathways in
Vibrio
cell (
Fig. S3A
), highlighted that only the AI-2 pathway was induced during nutrient stress of
V. atlanticus
LGP32, because only the expression of the AI synthase (
LuxS
) and its receptor (
LuxP
) increased significantly (
Fig. 4B
; ANOVA p <0.05). This was confirmed by a QS bioluminescence assay, which showed AI-2 molecules (unquantified) in the Zobell culture supernatant of
V. atlanticus
LGP32 after 60 h of culture but not after 12 h of culture and not in the ENSW supernatant of
V. atlanticus
LGP32 after 12 or 60 h of culture. UHPLC-HRMS/MS provided no evidence of detectable HAI-1 and CAI-1 in any experiments.
Targeted mutagenesis of key genes involved in two of the three known QS pathways in vibrios (
Fig. S3
), ΔluxM (HAI-1 production), ΔluxS (AI-2 production), and ΔluxR (main high-density QS regulator), did not result in any changes in the attack behavior of
V. atlanticus
LGP32 (
Fig. 4C
). Combined with the absence of overexpression of the CqsS gene (inducible by CAI-1) involved in the last known QS pathway in Vibrio (
Fig. S3
), these results indicated that the attack by
V. atlanticus
LGP32 is most likely unrelated to QS.
Attack related to the availability of iron
The comparative analysis of the proteome of
V. atlanticus
LGP32 incubated 60 h in artificial seawater (ENSW) versus
V. atlanticus
LGP32 grown 12 h in Zobell nutrient-rich medium revealed 10 proteins that were differentially abundant under these two contrasting conditions (
Fig. S2
). The two most down-regulated proteins correspond to be ß-ketoacyl-(acyl-carrier-protein) synthase II (-22-fold in ENSW compared to Zobell), a key regulator of bacterial fatty acid synthesis, and the dihydroorotase (-6.6-fold in ENSW compared to Zobell), an enzyme essential for pyrimidine biosynthesis and thus bacterial proliferation and growth. The low expression of these proteins in ENSW is consistent with
V. atlanticus
LGP32 nutritional starvation. The most up-regulated protein in starved
V. atlanticus
LGP32, with an increase of more than 6-fold, was glucosamine-6-phosphate deaminase, an enzyme involved in bacterial energy metabolism probably necessary for its survival. Among the other up-regulated proteins, one was an iron siderophore-binding protein (Spot 4413,
Fig. S2A
) corresponding to the vibrioferrin outer membrane receptor PvuB, whose gene is part of the
pvu
operons involved in iron transport (
Fig. S3B
). Interestingly, the corresponding gene
pvuB
as well as the vibrioferrin membrane receptor gene
pvuA2
(
Fig. S3B
) were both significantly induced in Vibrio under nutrient stress (
Fig. 4B
; ANOVA p <0.01) but not the one involved in the vibrioferrin biosynthesis,
pvsA
(
Fig. 4B
). Remarkably, of the 10 proteins identified by proteomic analysis and eliminated by mutation, only elimination of PvuB prevented
V. atlanticus
from attacking
A. pacificum
ACT03 (
Fig. 4C
; ANOVA p <0.001). In the absence of the
pvuB
gene,
V. atlanticus
LGP32 was unable to simultaneously attack
A. pacificum
ACT03. In addition,
V. atlanticus
LGP32 cells that had been washed with ENSW to remove their culture supernatant metabolites also failed to attack
A. pacificum
ACT03 (
Fig. 4C
; ANOVA p <0.001), which is congruent with the hypothesis that attacks depend on the
V. atlanticus
LGP32 vibrioferrin transport system. Finally, attacks increased significantly when FeCl
3
was added to the Vibrio culture medium (
Fig. 4D
) but not with boron known to be capable of being transported by vibrioferrin (see
Fig. 4D
). Taken together, those results are consistent with the hypothesis that attacks are regulated by iron.
Attack is a
Vibrio spp.
behaviour specific to
Alexandrium spp
To evaluate the dinoflagellates specificity of the attack behaviour, a selection of
Vibrio spp.
was co-cultured with a selection of dinoflagellate strains commonly found in the Mediterranean Sea. The results showed that, among the
Vibrio spp
. tested (pathogenic or not) all, when under nutrient stress, were able to secrete algicidal compounds, immobilize, attack and lyse
A. pacificum
ACT03 cells (
Table 1
) and no link to their pathogenicity for fish of invertebrates was observed (
Table 1
). Among the sixteen dinoflagellates species tested, only
Alexandrium spp
. (non-toxic and paralytic shellfish toxin (PST) producers) and
Gymnodinium catenatum
(PST producer) were immobilized, attacked and lysed by
V. atlanticus
LGP32 (
Table 2
), but no link to PSTs was revealed (
Table 2
).
Discussion
Predation is a widespread mode of interaction for survival in the natural world (
Finke and Denno, 2004
;
Sinclair et al., 2003
). Predatory bacteria are found in a wide variety of environments and are commonly described as feeding on other bacteria, although some cases of predation on microbial eukaryotes have also been hypothesized (
Johnke et al., 2014
;
Perez et al., 2016
). Conceiving predators as free-living organisms that kill other organisms and feed on them, this study suggests that Vibrio engage in a novel form of predation in which they kill and feed on algae.
In fact, the strategy developed by Vibrio to kill algae may be reminiscent of strategies previously described in the prokaryotes (
Johnke et al., 2014
). As shown in
Video 1
, the interaction between
V. atlanticus
LGP32 and
A. pacificum
ACT03 proceeds in three stages (
Fig. 5
). The first stage, the ‘
immobilization stage
’, recalls the strategy used by
Streptomyces
to immobilize its prey (
Kumbhar et al., 2014
) based on the secretion of algicidal metabolites that disrupt the flagella. The second stage, the ‘attack stage’ corresponding to the physical contact between Vibrios and Alexandrium, is similar to the strategy used by
Myxococcus xanthus
and Lysobacter. These bacteria may require close proximity to their prey to cause lysis and utilize their biomass, although some can also kill prey at a distance through diffusible secretions (
Martin, 2002
;
Genovesi et al., 2013
;
Perez et al., 2016
;
Zhang et al., 2020
).
V. atlanticus
LGP32 also surrounds
A. pacificum
ACT03 cells at high density for a very short time, but does not invade the algal cell. Visually, this phenomenon resembles bacteria clustering around lysed ciliate cells (
Blackburn et al., 1998
). The third stage, the ‘
killing stage
’, is similar to that of epibiotic bacterial predators, which induce the lysis of bacteria from the outside (
Rashidan and Bird, 2001
). Overall, these observations suggest that
V. atlanticus
LGP32 can exhibit a predatory behavior.
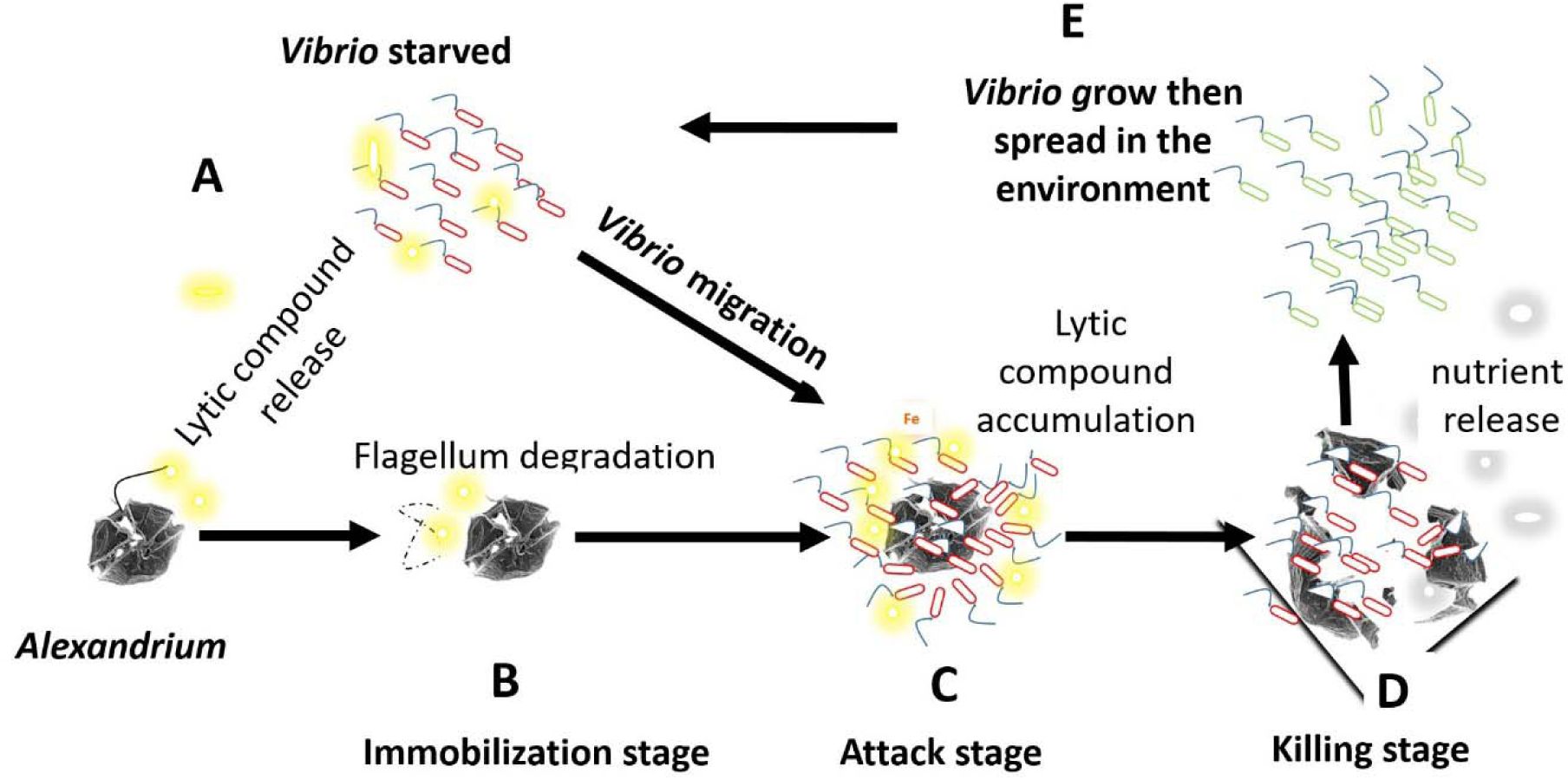
Schematic representation of a putative strategy developed by
Vibrio spp
. to feed on Alexandrium
spp
. and
G. catenatum
in the environment.
(A)
Vibrio in the environment when subjected to starvation secrete non-protein lytic compounds.
(B)
Some of these lytic compounds degrade the flagella, immobilizing the alga (immobilization stage).
(C)
Then Vibrio swims and clusters around its prey (attack stage).
(D)
Lytic compounds released by Vibrio where able to concentrate around the algal cells, thereby lysing the algae (killing stage).
(E)
Feeding on the released nutrients, Vibrio multiply and then spread in the environment. Yellow clouds: Lytic compound release by Vibrio, Grey clouds: Algal nutrients released upon lysis.
The attack behaviour of
V. atlanticus
LGP32 was linked to iron absorption, as mutants with impaired iron absorption completely lost its ability to attack the algae. Iron is an essential element for growth in most organisms, including phytoplankton (
Martin and Fitzwater, 1988
) and bacteria (
Neilands, 1981
), and its concentration in seawater is known to be very low with measurements in the open ocean surface at 0.2 nM and in deep waters at 0.6 nM (
Millero, 1998
).
Moreover, its low solubility in seawater limits its availability (
Bruland et al., 1994
;
Wu and Luther, 1994
). To acquire iron, bacteria have developed systems based on the secretion (and subsequent uptake) of iron-chelating siderophores to obtain this element from the environment (
Amin et al., 2009a
). Therefore, many
Vibrio spp
. produce a siderophore known as vibrioferrin, which is synthesized and secreted by proteins encoded by the
pvsABCDE
gene cluster (
Fig. S3
). Given that boron is known for its role in regulating a global bacterial cellular response to phytoplankton and to bind to vibrioferrin (
Romano et al., 2013
;
Weerasinghe et al., 2013
), we tested its potential involvement in simultaneous Vibrio attacks. Compared to the Zobell control, no effect on the number of attacks was observed. For iron-vibrioferrin uptake,
Vibrio parahaemolyticus
uses a membrane siderophore receptor, called PvuA, which is coupled to an inner membrane ATP-binding cassette (ABC). This ABC transporter system, which is comprised of proteins encoded by the pvuABCDE gene cluster (
Fig. S3
), is required for the transport of the siderophore across the inner membrane (
Tanabe et al., 2003
). Siderophores are not only iron carriers but also important regulators of virulence (
Miethke and Marahiel, 2007
) and mediators of bacterial interaction with phytoplankton (
Amin et al., 2009b
;
Kramer et al., 2020
). We showed here a pivotal role of iron in the interaction between
V. atlanticus
LGP32 and
A. pacificum
ACT03. This mirrors the mutualistic interaction observed between
Gymnodinium catenatum
and
Marinobacter
(
Amin et al., 2009b
). In fact, in natural settings, the co-occurrence of
Marinobacter
and
G. catenatum
is suggested to depend on a mutually beneficial utilization of iron and carbon resources (
Bolch et al., 2011
). As in the present study, iron seems to play a key role in the interaction. Indeed, the labile iron released through the photolysis of ferric chelates with vibrioferrin providing a crucial iron source for phytoplankton, which need substantial amounts of iron to support carbon fixation through photosynthesis (
Amin et al., 2009a
;
Yang et al., 2021
). This fixed carbon, in turn, sustains the growth of both the phytoplankton and their associated bacterial counterparts (
Amin et al., 2009b
;
Kramer et al., 2020
). Interestingly, if a general nutrient deficiency causes attacks, iron supplementation increases this number of attacks (
Figure 4D
), suggesting the importance of iron absorption in the attack behavior. Future studies should determine whether a nutrient deficiency increases the iron absorption capacity of Vibrio bacteria and whether this could play a major role in the attack mechanism.
This study showed that quorum sensing is not involved in microbial attacks. Thus,
V. atlanticus
LGP32 mutants lacking the genes involved in known quorum sensing pathways exhibit the same phenotype as wild-type
V. atlanticus
LGP32, and Vibrio density does not induce attacks. However, V. atlanticus LGP32 produces AI-2 during attacks. Quorum sensing and iron acquisition are sometimes interconnected in Vibrio (
McRose et al., 2018
). For example, in
Vibrio vulnificus
, the production of vulnibactin (a siderophore) is known to be controlled by AI-2 (
Kim and Shin, 2011
). Similarly, AI-2 could be involved in the production of vibrioferrin in
V. atlanticus
LGP32.
In the natural environment, associations between bacteria and algae have already been observed (
Lopez-Joven et al., 2018
;
Miller et al., 2005
;
Rosales et al., 2022
;
Xu et al., 2022
). We have shown here that algal attacks by Vibrio can be carried out
in vitro
. Environmental data collected in the Thau lagoon showed a correlation between the presence of Alexandrium and that of Vibrio, suggesting that such interactions could also occur in the marine environment. If that were the case, this behaviour would provide an important ecological advantage to Vibrio to obtain nutrients in the environment, where
Alexandrium spp
. and
Gymnodinium catenatum
form blooms.
With more than 30 species distributed all over the world (
Anderson et al., 2012
;
Hallegraeff et al., 2021
),
Alexandrium spp.
and
Gymnodinium spp
. considered as invasive species by the Delivering Alien Invasive Species Inventories for Europe (
)), could play an unexpected and important role in maintaining, structuring and regulating Vibrio populations in the ecosystem. In turn, Vibrio could contribute to the regulation and control of their blooms.
To conclude, this study reveals the capacity of some
Vibrio spp
. to act as facultative predator-like bacteria that hunt specific algae. In the current context of climate change, which is favourable to their development, monitoring the invasive algae
Alexandrium spp
. and
Gymnodinium catenatum
should be considered not only for their potent harmful effect on humans and animals, but also because they may represent a potential source of nutrients for the expansion of Vibrio, particularly pathogenic species (
Lemire et al., 2015
).
Figure supplements
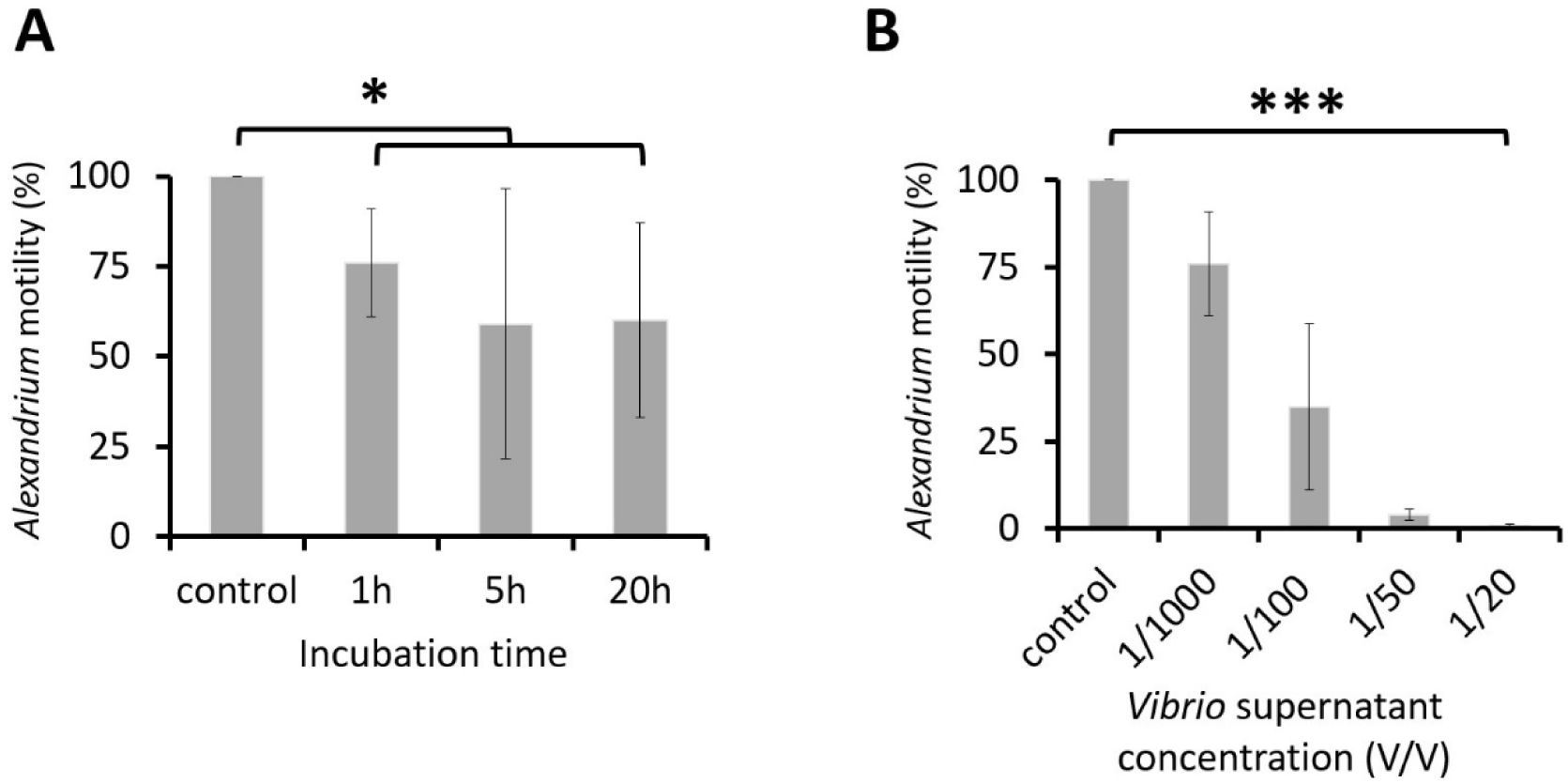
Time and dose-dependent effects of the
V. atlanticus
LGP32 culture supernatant on
A. pacificum
ACT03 motility.
(A)
A time dependence experiment was conducted by incubating
A. pacificum
ACT03 for 1, 5 or 20 h with 1/1000 v/v (1 µL/mL) of culture supernatant from
V. atlanticus
LGP32 previously grown for 60 h in Zobell culture media.
(B)
A dose-dependence experiment was conducted by incubating
A. pacificum
ACT03 for 1 h with 1/1000 to 1/20 v/v (1-50 µL/mL) of culture supernatant from
V. atlanticus
previously grown for 60 h in Zobell media. The percentage of motile
A. pacificum
ACT03 was determined after 1 hour of exposure. All percentages were determined based on a minimum of 2,000 cells of
A. pacificum
ACT03. Error bars represent the standard deviation of the mean of three independent experiments. Asterisks indicate significant differences in a multiple comparison test (One-way ANOVA with post hoc Tukey test), *P ≤ 0.05, ***P ≤ 0.001.

V. atlanticus
LGP32 proteome analysis following nutrient stress.
(A)
Example of 2D gel, the numbers in white on the gel 4-7 correspond to the number and position of the protein spots analyzed.
(B)
Proteins identified by LC-MS/MS as differentially represented in the 2D gel comparative approach following nutrient stress. ND: Not determined; ENSW (artificial seawater).
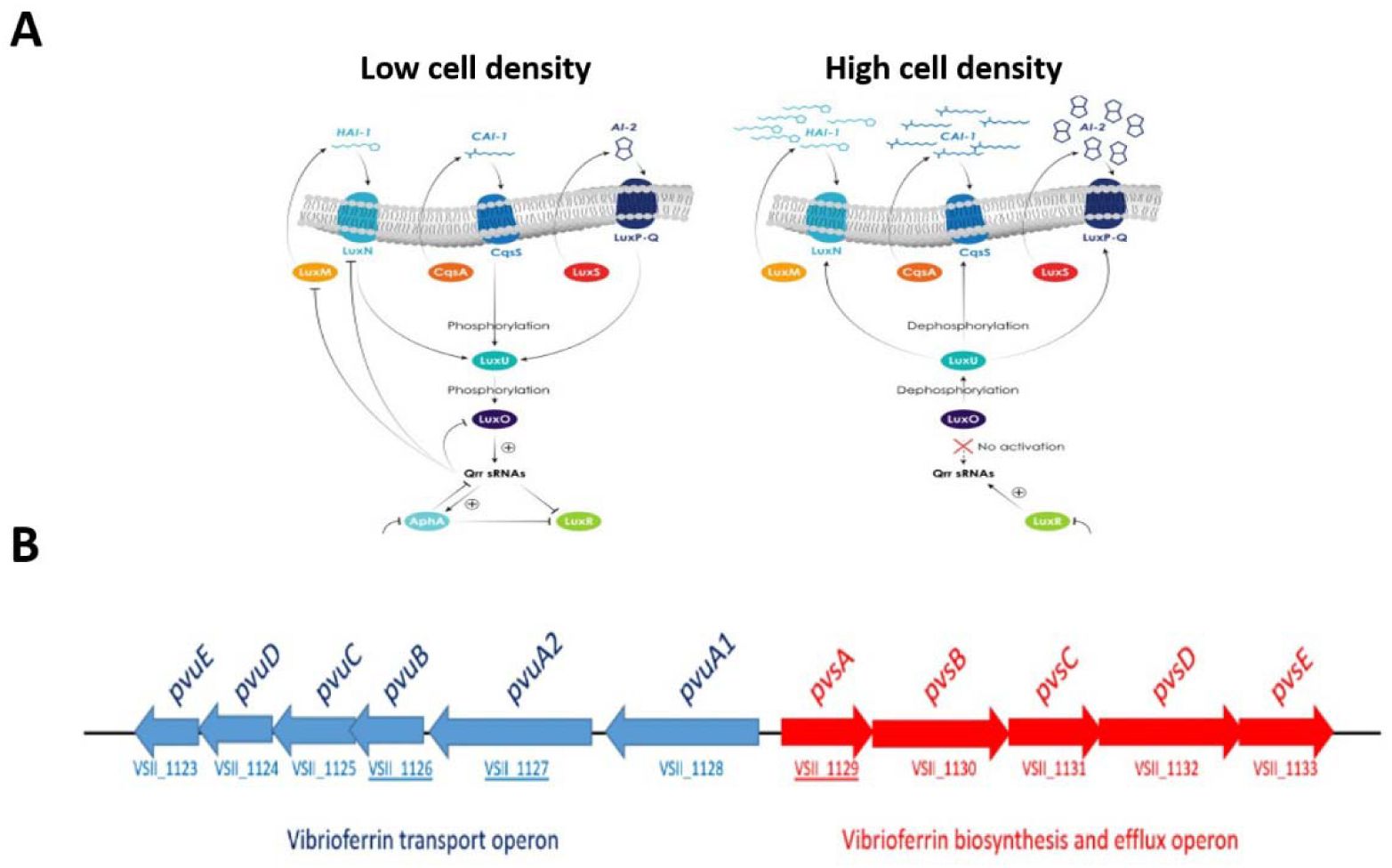
Quorum sensing and the vibrioferrin iron uptake pathway in Vibrio.
(A)
Putative quorum sensing (QS) pathways at low and high cell density in Vibrio according to Lami et al.(
Lami, 2019
).
(B)
Genetic organization of the vibrioferrin utilization gene cluster on
V. atlanticus
LGP32 chromosome 2. The Pvu and Pvs operons are involved in the secretion and the transport of ferric vibrioferrin and biosynthesis of vibrioferrin, respectively. Arrows indicate the transcriptional directions of the genes. VSII1126, VSII1137 and VSII1129 corresponding to PvuB, PvuA2 and PvsA genes respectively.
Data availability
Data generated or analysed during this study are included in the manuscript (tables, supplementary materials, videos).
Acknowledgements
The authors thank F. Le Roux and Y. Labreuche for their gift of
Vibrio
mutants; E. Garcés, S. Turki, H. Frehi, A. Bouquet and W. Medhioub for their contribution in obtaining Mediterranean phytoplankton strains; K. Escoubeyrou and D. Stien for advice on HRMS/MS data; V. Diakou-Verdin and E. Jublanc from the Montpellier RIO Imaging Platform for access to microscope facilities (
); P. Clair from the Montpellier qPHD platform and the Technoviv platform of the UPVD for access to PCR facilities; Luc Markiw, N. Brunet, A. Lang, A. Payelleville, T. Milhau and M. Leroy for their technical assistance; The IFREMER LER-LR laboratory for access to the Thau Lagoon; the BIO2MAR platform at the Observatoire Océanologique de Banyuls for providing access to UHPLC-HRMS/MS facilities.
Additional information
Funding
European VIVALDI grant 678589, French EC2CO ROSEOCOM grant and IFREMER ALGOVIR grant.
Funding
EC | Horizon 2020 Framework Programme (H2020)
Jean Luc Rolland
CNRS | Institut national des sciences de l'Univers (INSU,CNRS) (French EC2CO ROSEOCOM)
Jean Luc Rolland
Institut Français de Recherche pour l'Exploitation de la Mer (IFREMER) (ALGOVIR)
Jean Luc Rolland
Additional files
Video 1.
Dynamics of
V. atlanticus
LGP32
-Alexandrium pacificum
ACT03 interaction.
GFP-tagged
V. atlanticus
(small green cells); living
A. pacificum
(large red cells); lysed
A. pacificum
(large green cells) filmed under an epifluorescence microscope.
Video 2.
Second-by-second timing of
V. atlanticus
LGP32 attacking
Alexandrium pacificum
ACT03.
GFP-tagged
V. atlanticus
(small green cells);
A. pacificum
living cell (large red cells) filmed under an epifluorescence microscope.
Video 3.
Degradation and disruption of
Alexandrium pacificum
ACT03 flagella.
Effect of Vibrio supernatant on the first stage of the interaction filmed under a confocal microscope.
Video 4.
Attacks of
V. atlanticus
LGP32 on target
Alexandrium pacificum
ACT03.
This video, recorded under a confocal microscope, shows Vibrios simultaneously attacking a first immobilized Alexandrium cell, then moving on to attack a second cell without ever targeting the other cells present, suggesting active communication between the Vibrio bacteria.
V. atlanticus
LGP32 (small cells);
A. pacificum
ACT03 (large cells).
Video 5.
Vesicle formation and bursting of an
Alexandrium pacificum
ACT03.
Direct effect of Vibrio supernatant on
Alexandrium
after 126 h of culture filmed under a confocal microscope.
References
Contamination de l’étang de Thau par Alexandrium tamarense (No. Rapport 884)
Ifremer
Google Scholar
Effect of Nitrate, Ammonium and Urea on Growth and Pinnatoxin G Production of Vulcanodinium rugosum
Mar Drugs
13
:5642–5656
PubMed
Google Scholar
Photolysis of iron-siderophore chelates promotes bacterial-algal mutualism
Proc. Natl. Acad. Sci. U. S. A
106
:17071–17076
PubMed
Google Scholar
Vibrioferrin, an Unusual Marine Siderophore: Iron Binding, Photochemistry, and Biological Implications
Inorg Chem
48
:11451–11458
PubMed
Google Scholar
The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health
Harmful Algae
14
:10–35
PubMed
Google Scholar
Non-Cholera Vibrios: The Microbial Barometer of Climate Change
Trends Microbiol
25
:76–84
PubMed
Google Scholar
Toxicity and Growth Assessments of Three Thermophilic Benthic Dinoflagellates (Ostreopsis cf. ovata, Prorocentrum lima and Coolia monotis) Developing in the Southern Mediterranean Basin
Toxins
8
:297
PubMed
Google Scholar
Microscale nutrient patches in planktonic habitats shown by chemotactic bacteria
Science
282
:2254–2256
PubMed
Google Scholar
The toxic dinoflagellate gymnodinium catenatum (dinophyceae) requires marine bacteria for growth1
J. Phycol
47
:1009–1022
PubMed
Google Scholar
Reactive Trace-Metals in the Stratified Central North Pacific
Geochim Cosmochim Acta
58
:3171–3182
https://doi.org/10.1016/0016-7037(94)90044-2
Google Scholar
Environmental parameters associated with incidence and transmission of pathogenic Vibrio spp
Environ Microbiol
23
:7314–7340
PubMed
Google Scholar
Food Web and Ecosystem Impacts of Harmful Algae
In:
Shumway S.E.
Burkholder J.M.
Morton S.L
, editors.
Harmful Algal Blooms
John Wiley & Sons, Ltd
pp. 243–336
Google Scholar
Summary
In:
Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach
NY
:
Springer, New York
pp. 437–454
Google Scholar
Harmful algal blooms significantly reduce the resource use efficiency in a coastal plankton community
Sci. Total Environ
704
:135381
PubMed
Google Scholar
Algicidal Bacteria: A Review of Current Knowledge and Applications to Control Harmful Algal Blooms
Front. Microbiol
13
:871177
PubMed
Google Scholar
Interbacterial competition and anti-predatory behaviour of environmental Vibrio cholerae strains
Environ Microbiol
22
:4485–4504
PubMed
Google Scholar
The Impact of Protozoan Predation on the Pathogenicity of Vibrio cholerae
Front. Microbiol
11
:17
PubMed
Google Scholar
Alexandrium pacificum Litaker sp. nov (Group IV): Resting cyst distribution and toxin profile of vegetative cells in Bizerte Lagoon (Tunisia, Southern Mediterranean Sea)
Harmful Algae
48
:69–82
PubMed
Google Scholar
Predator diversity dampens trophic cascades
Nature
429
:407–410
PubMed
Google Scholar
Two Vibrio splendidus related strains collaborate to kill Crassostrea gigas: taxonomy and host alterations
Dis Aquat Organ
62
:65–74
PubMed
Google Scholar
Influences of sedimentation and hydrodynamics on the spatial distribution of Alexandrium catenella/tamarense resting cysts in a shellfish farming lagoon impacted by toxic blooms
Harmful Algae
25
:15–25
Google Scholar
Assessment of cryptic species diversity within blooms and cyst bank of the Alexandrium tamarense complex (Dinophyceae) in a Mediterranean lagoon facilitated by semi-multiplex PCR
J. Plankton Res
33
:405–414
Google Scholar
Unsuspected intraspecific variability in the toxin production, growth and morphology of the dinoflagellate Alexandrium pacificum R.W. Litaker (Group IV) blooming in a South Western Mediterranean marine ecosystem, Annaba Bay (Algeria)
Toxicon
180
:79–88
PubMed
Google Scholar
Perceived global increase in algal blooms is attributable to intensified monitoring and emerging bloom impacts
Commun. Earth Environ
2
:117
PubMed
Google Scholar
A Broad-Spectrum Artificial Seawater Medium for Coastal and Open Ocean Phytoplankton
J. Phycol
16
:28–35
Google Scholar
Multiple micro-predators controlling bacterial communities in the environment
Curr. Opin. Biotechnol
27
:185–190
PubMed
Google Scholar
Fitness Factors in Vibrios: a Mini-review
Microb Ecol
65
:826–851
PubMed
Google Scholar
Modulation of Iron-Uptake Systems by a Mutation of luxS Encoding an Autoinducer-2 Synthase in Vibrio vulnificus
Biol. Pharm. Bull
34
:632–637
PubMed
Google Scholar
Phylogenetic-Relationships of Marine-Bacteria, Mainly Members of the Family Vibrionaceae, Determined on the Basis of 16s Ribosomal-Rna Sequences
Int. J. Syst. Bacteriol
43
:8–19
PubMed
Google Scholar
Bacterial siderophores in community and host interactions
Nat. Rev. Microbiol
18
:152–163
PubMed
Google Scholar
Widespread predatory abilities in the genus Streptomyces
Arch. Microbiol
196
:235–248
PubMed
Google Scholar
Influence of temperature, salinity and irradiance on the growth and cell yield of the harmful red tide dinoflagellate Alexandrium catenella colonizing Mediterranean waters
J. Plankton Res
33
:1550–1563
Google Scholar
Wind and temperature controls on Alexandrium blooms (2000-2007) in Thau lagoon (Western Mediterranean)
Harmful Algae
28
:31–36
Google Scholar
Vibrio aestuarianus zinc metalloprotease causes lethality in the Pacific oyster Crassostrea gigas and impairs the host cellular immune defenses
Fish Shellfish Immunol
29
:753–758
PubMed
Google Scholar
Quorum Sensing in Marine Biofilms and Environments
In:
Tommonaro G
, editors.
Quorum Sensing
Academic Press
pp. 55–96
Google Scholar
Construction of a Vibrio splendidus Mutant Lacking the Metalloprotease Gene vsm by Use of a Novel Counterselectable Suicide Vector
Appl Environ Microbiol
73
:777–784
PubMed
Google Scholar
Seasonal variations of phytoplankton assemblages in relation to environmental factors in Mediterranean coastal waters of Morocco, a focus on HABs species
Harmful Algae
96
:101819
PubMed
Google Scholar
Populations, not clones, are the unit of vibrio pathogenesis in naturally infected oysters
Isme J
9
:1523–1531
PubMed
Google Scholar
The emergence of Vibrio pathogens in Europe: ecology, evolution, and pathogenesis (Paris, 11-12th March 2015)
Front. Microbiol
6
:830
PubMed
Google Scholar
A Novel Algicide: Evidence of the Effect of a Fatty Acid Compound from the Marine Bacterium, Vibrio sp BS02 on the Harmful Dinoflagellate, Alexandrium tamarense
Plos One
9
:e91201
PubMed
Google Scholar
News & Notes: Isolation of Vibrio harveyi from Diseased Kuruma Prawns Penaeus japonicus
Curr. Microbiol
33
:129–132
PubMed
Google Scholar
Oyster Farming, Temperature, and Plankton Influence the Dynamics of Pathogenic Vibrios in the Thau Lagoon
Front. Microbiol
9
:2530
PubMed
Google Scholar
Comparative genomics-based investigation of resequencing targets in Vibrio fischeri: Focus on point miscalls and artefactual expansions
Bmc Genomics
9
:138
PubMed
Google Scholar
Mediterranean alien harmful algal blooms: origins and impacts
Environ Sci Pollut Res
28
:3837–3851
PubMed
Google Scholar
Iron-Deficiency Limits Phytoplankton Growth in the Northeast Pacific Subarctic
Nature
331
:341–343
Google Scholar
Predatory prokaryotes: An emerging research opportunity
J. Mol. Microbiol. Biotechnol
4
:467–477
PubMed
Google Scholar
Toxigenic Vibrio cholerae evolution and establishment of reservoirs in aquatic ecosystems
Proc. Natl. Acad. Sci
117
:7897–7904
PubMed
Google Scholar
Quorum sensing and iron regulate a two-for-one siderophore gene cluster in Vibrio harveyi
Proc. Natl. Acad. Sci
115
:7581–7586
PubMed
Google Scholar
Siderophore-Based Iron Acquisition and Pathogen Control
Microbiol. Mol. Biol. Rev
71
:413–451
PubMed
Google Scholar
Detection of a bioluminescent milky sea from space
Proc. Natl. Acad. Sci. U. S. A
102
:14181–14184
PubMed
Google Scholar
Solubility of Fe(III) in seawater
Earth Planet Sci Lett
154
:323–329
https://doi.org/10.1016/S0012-821X(97)00179-9
Google Scholar
Projections of the future occurrence, distribution, and seasonality of three Vibrio species in the Chesapeake Bay under a high-emission climate change scenario
Geohealth
1
:278–296
PubMed
Google Scholar
Iron-Absorption and Transport in Microorganisms
Annu. Rev. Nutr
1
:27–46
PubMed
Google Scholar
Seasonal Dynamics and Modeling of a Vibrio Community in Coastal Waters of the North Sea
Microb Ecol
63
:543–551
PubMed
Google Scholar
Biotic control of harmful algal blooms (HABs): A brief review
J. Environ. Manage
268
:110687
PubMed
Google Scholar
Quorum sensing signal-response systems in Gram-negative bacteria
Nat. Rev. Microbiol
14
:576–588
PubMed
Google Scholar
Bacterial predation: 75 years and counting! Environ
Microbiol
18
:766–779
PubMed
Google Scholar
A new mathematical model for relative quantification in real-time RT-PCR
Nucleic Acids Res
29
:e45
PubMed
Google Scholar
R: A Language and Environment for Statistical Computing
Role of predatory bacteria in the termination of a cyanobacterial bloom
Microb Ecol
41
:97–105
PubMed
Google Scholar
Straightforward N -Acyl Homoserine Lactone Discovery and Annotation by LC–MS/MS-based Molecular Networking
J. Proteome Res
21
:635–642
PubMed
Google Scholar
Paralytic Toxins Accumulation and Tissue Expression of alpha-Amylase and Lipase Genes in the Pacific Oyster Crassostrea gigas Fed with the Neurotoxic Dinoflagellate Alexandrium catenella
Mar Drugs
10
:2519–2534
PubMed
Google Scholar
Regulation of iron transport related genes by boron in the marine bacterium Marinobacter algicola DG893
Metallomics
5
:1025
PubMed
Google Scholar
Investigating the Relationship between Nitrate, Total Dissolved Nitrogen, and Phosphate with Abundance of Pathogenic Vibrios and Harmful Algal Blooms in Rehoboth Bay, Delaware
Appl Environ Microbiol
88
:e00356–22
PubMed
Google Scholar
Patterns of predation in a diverse predator-prey system
Nature
425
:288–290
PubMed
Google Scholar
Isolation and characterization of a marine algicidal bacterium against the toxic dinoflagellate Alexandrium tamarense
Harmful Algae
6
:799–810
Google Scholar
Identification and Characterization of Genes Required for Biosynthesis and Transport of the Siderophore Vibrioferrin in Vibrio parahaemolyticus
J. Bacteriol
185
:6938–6949
PubMed
Google Scholar
Vibrio tasmaniensis sp. nov., isolated from Atlantic Salmon (Salmo salar L.)
Syst Appl Microbiol
26
:65–69
PubMed
Google Scholar
Bacterial–Fungal Interactions in the Kelp Endomicrobiota Drive Autoinducer-2 Quorum Sensing
Front. Microbiol
10
:1693
PubMed
Google Scholar
Copper homeostasis at the host vibrio interface: lessons from intracellular vibrio transcriptomics
Environ Microbiol
18
:875–888
PubMed
Google Scholar
Lysis of a red-tide causing alga, Alexandrium tamarense, caused by bacteria from its phycosphere
Biol. Control
52
:123–130
Google Scholar
Continuous production of algicidal compounds against Akashiwo sanguinea via a Vibrio sp. co-culture
Bioresour Technol
295
:122246
PubMed
Google Scholar
Borate as a Synergistic Anion for Marinobacter algicola Ferric Binding Protein, FbpA: A Role for Boron in Iron Transport in Marine Life
J. Am. Chem. Soc
135
:14504–14507
PubMed
Google Scholar
Size-Fractionated Iron Concentrations in the Water Column of the Western North-Atlantic Ocean
Limnol Oceanogr
39
:1119–1129
Google Scholar
Co-occurrence of chromophytic phytoplankton and the Vibrio community during Phaeocystis globosa blooms in the Beibu Gulf
Sci. Total Environ
805
:150303
PubMed
Google Scholar
Marinobacter alexandrii sp. nov., a novel yellow-pigmented and algae growth-promoting bacterium isolated from marine phycosphere microbiota
Antonie Van Leeuwenhoek Int J Gen Mol Microbiol
114
:709–718
PubMed
Google Scholar
Dynamics of Solitary Predation by Myxococcus xanthus on Escherichia coli Observed at the Single-Cell Level
Appl Environ Microbiol
86
:e02286–19
PubMed
Google Scholar
Article and author information
Author information
Version history
Preprint posted
:
Sent for peer review
:
Reviewed Preprint version 1
:
Reviewed Preprint version 2
:
Reviewed Preprint version 3
:
Cite all versions
You can cite all versions using the DOI
10.7554/eLife.107221
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2025,
Rolland et al.
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
views
0
downloads
27
citations
0
Views, downloads and citations are aggregated across all versions of this paper published by eLife.





