 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库SCLS | 中科大张凯铭/李珊珊团队与龚庆国团队联合解析RNA甲基转移酶NSUN2识别与重塑底物tRNA的分子机制


RNA修饰被誉为生命活动中的“隐形密码”,通过在RNA分子上添加特定的化学标记,精细调控着基因表达与细胞命运。在真核生物广泛存在的众多RNA修饰中,5-甲基胞嘧啶(m⁵C)修饰占据着重要地位,它不仅参与基因表达调控,更与神经发育、应激响应以及肿瘤的发生发展密切相关。然而,作为催化该修饰的核心酶,NSUN2是如何在众多RNA分子中准确识别靶标并完成修饰的,一直是RNA生物学领域亟待解决的关键科学问题。

2026年5月26日,Science China Life Sciences (《中国科学:生命科学》英文版)在线发表中国科学技术大学张凯铭/李珊珊团队与龚庆国团队合作撰写的题为“Structure-driven RNA remodeling underlies broad substrate recognition by NSUN2”的研究论文,该工作综合运用X射线晶体学、单颗粒冷冻电子显微镜以及生化分析等技术,成功解析了人源RNA甲基转移酶NSUN2在无底物状态,以及结合不同tRNA底物状态下的高分辨率三维结构,揭示了NSUN2通过主动重塑RNA结构实现广谱底物识别的分子机制。
值得关注的是,5月27日,国际同行也在Nature上报道了NSUN2底物选择性的相关结构研究,进一步凸显了该领域受到的广泛关注。
双结构域协同:NSUN2对tRNA的锚定与捕获
在NSUN家族中,NSUN2拥有广泛的底物谱系(包括成熟tRNA、前体tRNA以及mRNA等),但其广谱识别的结构基础长期悬而未决。为此,研究团队利用催化位点突变体,成功捕获了NSUN2与多种不同tRNA底物(tRNATyr、tRNALys以及pre-tRNALeu)形成的稳定中间态复合物。结构分析表明,NSUN2主要依赖其N端的MTase结构域与C端的PUA-like结构域协同完成对底物的识别:前者负责碱基识别和催化,后者负责锚定tRNA并稳定结合。值得注意的是,当NSUN2结合tRNA后,其催化中心附近的螺旋区域会向内靠近,形成更加紧凑识别口袋,同时多个关键催化残基侧链重排调整取向,向活性中心靠近,为后续甲基化反应做足准备。
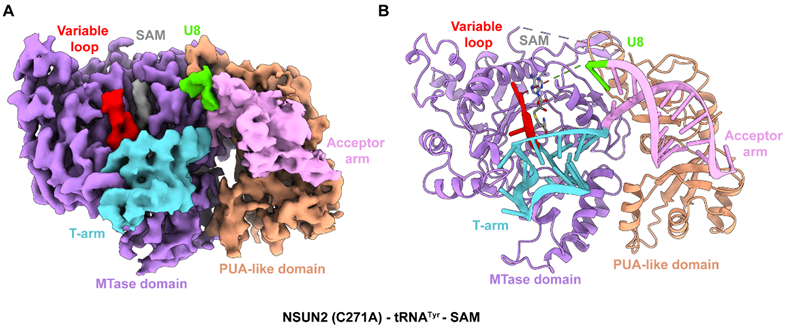
图1: NSUN2-tRNATyr-SAM 三元复合物的冷冻电子显微镜结构
结构驱动型识别:主动重塑tRNA构象
在天然状态下,tRNA呈现经典的“L”形三维结构,其修饰靶位点C48深埋于RNA内部。研究团队发现,NSUN2并非被动等待靶位点暴露,而是主动驱动底物发生大规模构象重排:促使tRNA的T臂向外翻转,由闭合构象转变为更加开放的构象,破坏原有的水平堆积作用;进一步促使原本埋藏的C48发生“碱基翻转”并暴露出来,精准送入NSUN2的催化中心。生化实验进一步证实,即使破坏了维持tRNA经典L形构象的关键相互作用,NSUN2仍能正常结合并修饰tRNA。这一结果证明了该酶的识别过程更依赖于底物的局部结构特征,而非完整的L形构象,从而揭示了一种由酶自身保持相对稳定、却主动诱导RNA发生适应性构象变化的“结构驱动型”精准修饰模式。
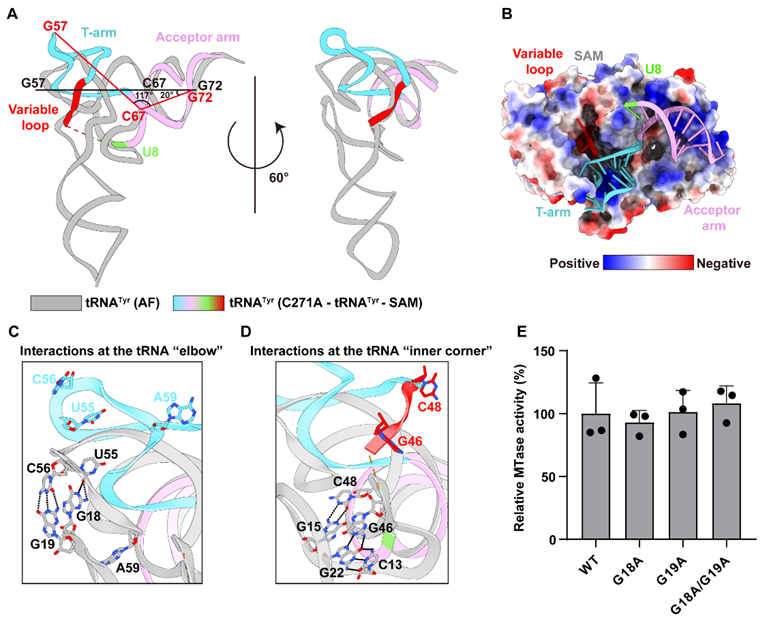
图2: tRNAᵀʸʳ在识别过程中的构象变化。A. AlphaFold 3预测tRNAᵀʸʳ结构与NSUN2-tRNAᵀʸʳ-SAM复合物中观测结构的对比。B: NSUN2与tRNAᵀʸʳ的结合界面。tRNAᵀʸʳ以棍状模型表示,NSUN2以静电势表面表示。C-D. AlphaFold 3预测与复合物观测结构中,tRNAᵀʸʳ肘部和内角相互作用的细节视图。E. 体外纯化的NSUN2对野生型及突变体tRNA的催化活性检测。数据以平均值±标准差(n = 3次独立实验)表示
广谱tRNA底物识别机制:依赖保守tRNA结构特征而非严格序列
通过对比NSUN2与多种不同tRNA底物的复合物结构,研究团队发现,尽管这些tRNA在序列、长度上存在明显差异,但它们与NSUN2结合时均采用高度相似的构象和结合方式。NSUN2的结合界面主要聚焦于tRNA所共有的受体臂、T臂和可变环等保守结构元件,而非依赖于严格的序列基序。这种对共有拓扑结构的普适性识别,为NSUN2广谱底物识别能力提供了合理解释。
基于上述“结构驱动”的普适规律,研究者成功设计并构建了最小化的“小型类tRNA(small tRNA-mimic)”底物。实验结果表明,仅保留受体臂、T臂及可变环,即可维持高效的甲基化反应,优化后的体系催化活性高达野生型的88%左右。这一工程化设计不仅阐明了NSUN2的保守相互作用界面,也暗示了其修饰具有相似局部结构的mRNA和lncRNA的潜在机制。
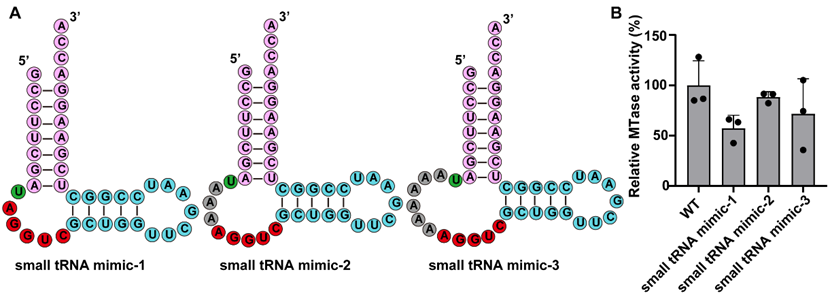
图3: 最小化“小型类tRNA”底物设计。A. 三种小型类tRNA的二级结构图。B. 纯化的NSUN2对野生型及不同小型类tRNA的催化活性检测。数据以平均值±标准差(n = 3次独立实验)表示
明确pre-tRNA的“首选修饰位点”
有趣的是,尽管此前有报道称NSUN2主要负责前体tRNALeu第34位点的修饰,但该研究的高分辨率冷冻电镜结构清晰显示,C48位点是其在复合体中的主要占据点。后续生化实验进一步证实,虽然NSUN2兼具修饰两个位点的能力,但在催化效率上对C48位点表现出了更为显著的修饰偏好性。
综上,该研究通过捕捉NSUN2在催化前后的不同动态,建立底物识别与催化模型。这一发现解释了NSUN2如何通过识别tRNA的保守结构特征而非特定序列来维持广谱的底物修饰能力。该成果不仅完善了对NSUN家族酶促机制的理解,也为未来针对NSUN2相关恶性肿瘤等疾病的靶向药物研发提供了坚实的结构生物学基础。
Science China Life Sciences同期发表复旦大学生命科学学院麻锦彪教授撰写的题为“How NSUN2 reshapes tRNA for m5C modification”的Research Highlight文章,对该研究进行了评论与解读:认为该研究为理解NSUN2广谱底物识别机制提供了重要结构基础,并推动了RNA修饰酶“结构驱动识别”理论框架的发展。
中国科学技术大学生医部博士后胡倩、杨雯、博士研究生余云云和伊然为该文的共同第一作者;硕士研究生张雨彤和博士研究生段乐园亦为研究做出了重要贡献。李珊珊副教授、龚庆国教授和张凯铭教授为该文共同通讯作者。
该研究工作获得了国家自然科学基金、国家重点研发计划、中国科学院战略性先导科技专项,以及合肥综合性国家科学中心大健康研究院等项目的资助。研究同时得到了中国科学技术大学、华中科技大学和安徽医科大学第一附属医院冷冻电镜中心以及中国科学技术大学附属第一医院的大力支持。
[点击下方链接或阅读原文]▼
https://doi.org/10.1007/s11427-026-3373-3
专题|RNA修饰:连接分子机制与临床应用的桥梁
深度综述|tRNA甲基化修饰及其在癌症中的作用:从分子机制到药物研发
核酸修饰驱动的分子医学:现状、挑战与机遇







