 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库Cell:冷冻电镜破解 VPS13A-XKR1 复合物结构,脂质“抄近道”直接跨膜转移
在真核细胞中,构成细胞器膜的脂质主要在内质网(ER)合成,然后被重新分配到其他细胞器。长期以来,人们知道细胞内膜之间的脂质转移可以通过囊泡运输实现。但近年认识到,桥状脂质转运蛋白(BLTP)在大量脂质
运动
(不依赖囊泡)中起着核心和基础作用。BLTP 通过“桥结构域”在紧密并列的细胞器之间跨越,桥结构域上有一条疏水沟槽,脂质可沿此沟槽在细胞器膜之间流动。
BLTP 介导的大量脂质转移被认为是伴随新细胞器(如自噬体)生物发生的膜扩展、囊泡运输途径之外细胞器(如线粒体)的膜维持,以及受损溶酶体膜的修复等过程的关键。BLTP 家族的创始成员 VPS13 蛋白也具有重要的生物医学意义,因为它们的
功能障碍
会导致神经疾病,如
帕金森病
(VPS13C)或舞蹈病-棘红细胞增多症(VPS13A);它们与自噬中起作用的 ATG2 家族一起,是研究最多的 BLTP。
一个新兴的范式(主要基于 VPS13 和 ATG2 蛋白的研究)是:在受体膜上,BLTP 与膜嵌入的 scramblase(一种能在膜双层之间平衡磷脂的蛋白)合作。这种合作使得尽管 BLTP 仅将脂质递送到膜的一个小叶,但膜的两个小叶可同时扩张。在缺乏 BLTP-scramblase 复合物近原子分辨率信息的情况下,控制脂质从桥结构域转移到膜以及 scramblase 在此过程中的作用机制一直难以捉摸。

在一项新的研究中,研究者结合结构和计算方法,研究了 VPS13 家族成员——人 VPS13A 如何与 XK 相关家族中的质膜 scramblase(称为 XK 或 XKR1)合作,以递送脂质。VPS13A 的功能丧失突变导致舞蹈病-棘红细胞增多症,这是一种亨廷顿病样的神经退行性疾病,伴有血液中异常红细胞(棘红细胞)。一种非常相似的疾病——McLeod 综合征,是由 XKR1 的功能丧失突变引起的,这与两者功能合作以及已报道的直接相互作用一致。
此外,VPS13A 和 XKR1 是在 T 细胞中磷脂酰丝氨酸表面暴露所需的基因筛选中排名最高的两个基因,提示一个模型:VPS13A 将磷脂从 ER 递送到质膜,然后 XKR1 将其翻转。VPS13A 也定位于细胞其他位置,如 ER 与线粒体之间的接触点,其功能研究较少。
研究者利用冷冻电镜(cryo-EM)以近原子分辨率(三个不同图谱,分辨率 3.28–3.41 Å)可视化 VPS13A、最近鉴别的可溶性相互作用伴侣钙调蛋白(CaM)以及去垢剂增溶的 XKR1 的复合物。VPS13A 与 XKR1 的相互作用将 VPS13A 的桥结构域定向在紧邻脂双层的位置,并将 VPS13A 固定在 scramblase 附近。分子动力学(MD)模拟表明,该冷冻电镜结构与桥结构域和受体膜的胞质小叶之间稳健的直接脂质转移相容,提示膜特性(如膜
张力
)可能将脂质递送与 XKR1 的脂质翻转偶联起来。研究者预期,此处描述的关于 VPS13A 脂质递送的机制见解可直接应用于所有 VPS13 和 ATG2 蛋白,以及更广泛的其它 BLTP 家族。
总之,VPS13A/CaM-XKR1 复合物的实验结构和 MD 分析表明,当 VPS13A 与其在受体膜中的结合伙伴 XKR1 对接时,脂质递送被激活,然后脂质直接从 VPS13A 的桥结构域转移到质膜的胞质小叶。VPS13A 的 PH 结构域与 XKR1 的相互作用、VPS13A 的 C 端帽和 VAB 结构域的构象变化,以及 VPS13A 的 ATG2_C 基序中双亲性螺旋的膜插入,对于将桥结构域的末端并置到近端膜是必需的,从而使脂质能够从桥内的疏水环境移动到膜中,而无需其酰基链去溶剂化。研究表明,VPS13A-XKR1 复合物在受体膜(预期发生 VPS13A 桥的脂质转移处)引入了显著的局部膜变形。这些变形影响脂质堆积,并可能降低脂质插入受体膜的能垒。
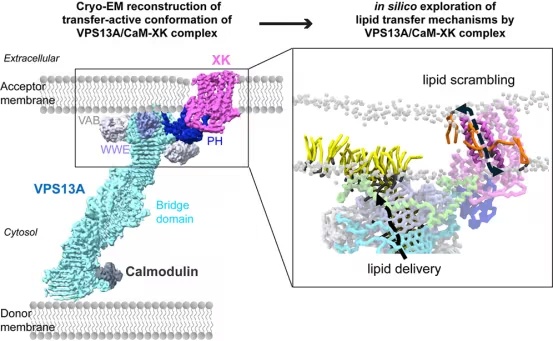
重要的是,当桥结构域内存在过量脂质(模拟 ER 中的脂质合成)时,研究者能够在计算机模拟中观察到稳健的脂质转移。此外,受体膜张力的增加加速了脂质释放。这些结果支持当前的观点:方向性的 BLTP 介导的脂质流可能由供体膜和受体膜之间的膜张力差异驱动。ER(大多数结构脂质合成的供体膜)具有低膜张力,而其他细胞器的膜张力较高。目前尚不清楚膜张力差异如何产生,这是一个备受关注的话题。
VPS13A 将其脂质递送到 XKR1 附近的膜的胞质小叶,然后 XKR1 可以将新递送的脂质在双层的两个小叶之间平衡。计算机评估表明,在冷冻电镜重建中观察到的 XKR1 构象不支持脂质翻转,其他 XK 家族成员的实验结构也不支持。然而,XKR1 的 AA-MD 模拟表明存在另一种构象,其中 TM1 和 TM4 分开形成一个极性通道,脂质可以通过该通道在双层小叶之间滑动。XKR1 和 XKR1-PH 融合构建体在体外嵌入脂质体时都能翻转脂质,支持 XKR1 可能在膜中采用一种翻转活性构象,而不是像冷冻电镜重建中那样在去垢剂胶束中。这一观察结果与质膜上的 TMEM16 scramblase 的观察结果相似,后者也通过亲水沟槽的开闭来调节脂质翻转活性。对 XKR4 和 XKR8 的突变分析与其一致,但 XK 相关家族的其他成员是否通过相同机制发挥作用仍有待研究。
有趣的是,计算机模拟表明,较高的膜张力既加速了 VPS13A 的脂质递送,又促进了 XKR1 的开放(即具有脂质翻转能力的构象)。总之,这些观察结果表明,尽管 VPS13A 的桥结构域和 XKR1 之间没有直接的物理连接,但 BLTP 和 scramblase 的脂质递送仍然可能通过双层特性的物理或化学修饰耦合。
在研究的 VPS13A-XKR1 复合物中,其 PH 结构域在将 VPS13A 靶向质膜上的 XKR1 中起主要作用,并与 ATG2_C 基序协调,将桥结构域牢固地锚定在膜上。不排除 VAB 和 WWE 结构域可能通过额外的相互作用辅助膜结合。此外,VPS13A 也与其他受体在细胞其他位置发挥作用,其中 VAB 或 WWE 结构域或 PH、VAB 和 WWE 结构域的某种组合可能介导细胞器附着。研究者设想,VAB 或 WWE 结构域可能具有与 PH 结构域相同的功能,即相对于受体膜定位桥结构域。包括 VPS13B、VPS13C 和 VPS13D 在内的人 VPS13 蛋白以及其它生物中的 VPS13 蛋白具有相似的结构,共享内置的衔接模块(ATG2_C、PH、VAB;WWE 结构域仅存在于 VPS13A 和 VPS13C 中)。因此,此处描述的 VPS13A/CaM-XKR1 复合物的结构可用于理解所有 VPS13 蛋白的脂质递送。
与 VPS13 相关的 ATG2 蛋白缺乏 VPS13 家族的大部分内置衔接结构域,而是依赖外源伙伴蛋白进行定位。它们是 VPS13 的精简版本,仅保留桥结构域和 ATG2_C 双亲性螺旋。在自噬过程中,ATG2 与 scramblase ATG9 合作,并可直接结合。基于当前对 VPS13A 脂质递送的研究,研究者提出:脂质从 ATG2 的桥结构域直接转移到膜,其中 ATG2_C 基序和 ATG2-ATG9 相互作用将桥结构域并置到受体膜附近,同时将 BLTP 定位在 scramblase 附近。最近的 MD 研究支持 ATG2 可以直接将脂质递送到受体膜,并描述了 ATG2_C 双亲性螺旋在此过程中膜变形中的作用。值得注意的是,在自噬过程中,ATG9 定位于新生自噬体的边缘,该边缘高度弯曲。
与本研究模型相反,早期对 ATG2-ATG9 复合物的低分辨率研究曾推测 ATG2 将脂质递送给 scramblase 而非直接给膜,然后 scramblase 将脂质整合到膜中。对早期数据的另一种解释与本研究提出的模型一致,即 ATG2 被拴系到 ATG9 上以将其带到靠近翻转活性的膜附近,同时脂质从桥结构域直接转移到膜。因此,本研究显著推进了人们对 ATG2 家族中 BLTP 脂质递送的理解。其他研究较少的 BLTP 家族(其成员也缺乏 ATG2_C 双亲性螺旋)也可能直接将脂质递送到受体膜,但负责膜结合和变形的特征仍有待鉴定。
最近一项关于形成于供体膜(ER)上的含 BLTP1 复合物的原子分辨率研究表明,脂质很可能通过一个由五个 TM 螺旋组成的整合膜蛋白前庭进入 BLTP1 桥结构域。本研究对 BLTP1 研究进行了补充,描述了脂质从桥结构域输出并返回到膜的机制。可以说出乎意料的是,受体膜上的脂质转移过程(至少对于 VPS13 而言,脂质直接转移到膜)并不反映在 BLTP1 上观察到的供体膜提取过程。研究者期待未来对 BLTP 蛋白及其两端过程的研究,以帮助理解每种策略的普遍性。
研究局限性
在冷冻电镜研究中,XKR1 被溶解在去垢剂胶束中而非嵌入膜中。因此,尽管结构显示 VPS13A 桥结构域的 C 端定位为与受体膜直接相互作用(基于 XKR1 结构推断了膜平面),但冷冻电镜数据并未直接显示 VPS13A 与膜的相互作用。即,从实验结构尚不清楚辅助结构域(如 VAB、WWE 或 PH 结构域)是否与膜相互作用,更重要的是,关于脂质转移过程,C 端帽和 ATG2_C 基序的双亲性螺旋相对于脂双层的排列如何。在缺乏可视化相互作用的实验结构的情况下,研究者依赖 MD 模拟来更好地理解复合物如何与膜对接以递送脂质。
MD 模拟有其固有的准确性局限性(取决于所使用的力场,全原子或粗粒度)和采样限制,在本研究通过执行多个微秒级的独立重复来缓解。对于粗粒度模拟,这可能导致过程加速和非物理速率,例如针对翻转和脂质递送测量的速率。对于粗粒度模拟,另一个局限性是使用弹性网络来维持冷冻电镜结构中发现的二级结构。然而,对于非结构或无序结构域(如 ATG2_C 基序)未使用弹性网络,它们可以根据力场能量景观自由采样其构象空间。
当应用于冷冻电镜结构时,MD 模拟支持桥结构域与膜之间直接转移脂质是合理的,特别是在存在“驱动力”(例如供体膜中的脂质生物合成)的情况下。在重构系统中测试计算机预测的一个局限性是尚未开发出具有生理相关性的体外试验。尽管研究者和其他人已经证明 BLTP 可以在脂质体之间转移脂质,但在这些体外系统中,BLTP 很可能作为脂质体之间的穿梭机。
迄今为止,该领域尚未重构出一个能证明 BLTP 作为脂质定向流动通道的系统。有两个主要障碍需要克服。首先,介导 VPS13 与供体膜(通常为 ER)结合的蛋白尚未完全确定,因此不清楚如何以生理相关方式将 VPS13 附着到供体膜。其次,尚未开发出重现膜张力增加的方案,正如供体脂质体仅在外小叶进行脂质生物合成可能产生的结果。研究者期待这类试验的开发,这对于评估驱动力的性质至关重要。(
生物谷)
参考文献:
Bodan Hu et al,
Mechanism of lipid transfer by bridge-like protein VPS13A and the scramblase XK
, Cell (2026). DOI: 10.1016/j.cell.2026.05.027.





