 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库中国药科大学孙建国:丹皮酚的药理作用及药代动力学研究进展
丹皮酚的药理作用及药代动力学研究进展 PPS
赵紫雅1,2,孙建国1,2,3*
(1. 中国药科大学 江苏省药物代谢动力学研究重点实验室,江苏 南京210009;2. 中国药科大学多靶标天然药物全国重点实验室,江苏南京 210009;3. 中国医学科学院“ 中药复杂组分PK-PD结合研究 ”创新单元,北京 100730)
孙建国

博士,研究员,博士研究生导师,专业方向为药物代谢动力学,主要从事创新药物体内和体外吸收及代谢特性研究、药物相互作用研究 、生理药代动力学研究、中药药代动力学研究 。先后赴新西兰奥塔哥大学药学院和美国华盛顿大学药学院任访问学者,2010 年赴新西兰参加科技部组织的“ 中新科学家交流计划”,2011 年被评为江苏省“333高层次人才培养工程 ”第三梯队培养对象并获得政府留学奖学金 。担任 J Chromatogr B, Xenobiotica, Phytomedicine与《药学进展》《中国新药杂志》等杂志审稿人。
先后主持完成了4项江苏省自然科学基金资助项目,2项国家自然科学基金资助项目,1项国家重点研发计划资助项目,主持多项国家重大科研项目子课题,参加多项国家重大科研项目,近年来发表论文40余篇,其中以第一作者或通信作者发表的论文20余篇(SCI论文18篇),申请专利6项,授权4项。曾获国家科技进步奖二等奖1项,全国创新争先团队奖1项,江苏省科技进步奖一等奖3项。与企业共同合作获得新药临床批件5项,负责完成的药代动力学研究支持企业获得新药证书多项。
[摘要] 牡丹皮具有清泄心、肾、肝之火以及活血化瘀等功效,在中医临床中应用广泛。丹皮酚作为牡丹皮的核心活性单体,因其多靶点生物活性成为天然药物研究热点。药理学研究表明,丹皮酚通过调控炎症介质、细胞周期蛋白及氧化应激通路,在抗炎、神经保护、心血管保护、抗肿瘤等方面展现出多维度治疗潜力, 同时其药代动力学特征也逐步被揭示。基于国内外相关研究,系统总结了丹皮酚的药理作用及药代动力学研究进展, 旨在为丹皮酚的进一步研究和临床应用提供理论依据和新的思路。
丹皮酚分离自牡丹、芍药的干燥根皮以及徐长卿的根或全草[1],是中药本草牡丹皮的主要活性成分。牡丹皮作为中国传统中药材,其应用历史可追溯至《神农本草经》和《本草纲目》,其中详细记载了牡丹皮清泄心、肾、肝之火以及活血化瘀的功效,并在经典方剂如仲景肾气丸中得到广泛应用。
随着现代科学技术的发展,牡丹皮的化学成分和药理作用研究取得了显著进展。丹皮酚作为牡丹皮中最重要的药效成分,已被证实具有广泛的药理活性[2-5](见图1)。本文总结丹皮酚在抗炎、神经保护、心血管保护、抗肿瘤等方面的药理作用与相关机制以及其在药代动力学方面的研究进展,以期为深入探究丹皮酚在临床应用中的潜在价值提供参考。
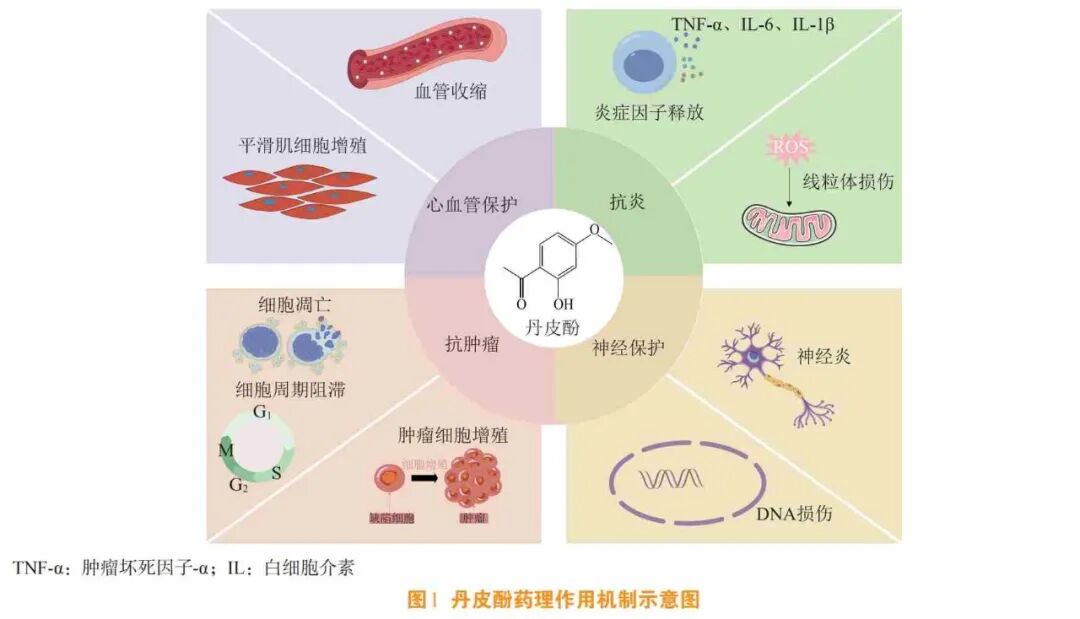
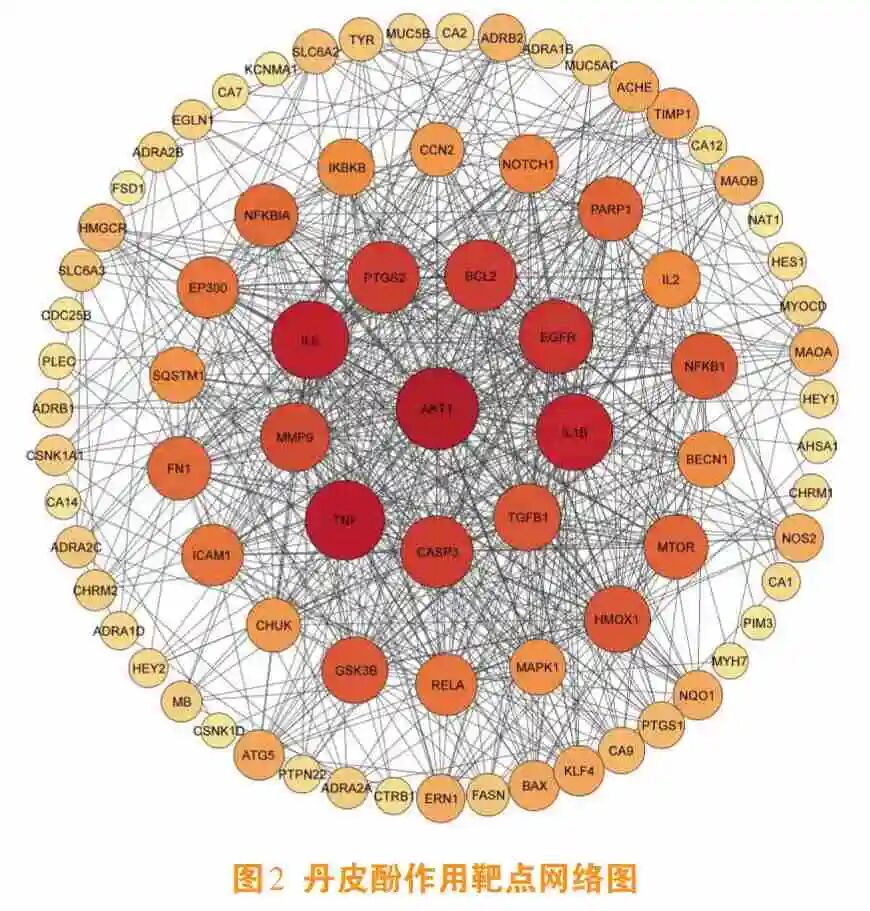
笔者所在课题组基于中医药整体调节理论,深入解析了丹皮酚的多靶点作用特征。通过整合TCMSP与CTD数据库资源,构建了丹皮酚的分子相互作用网络图谱,对丹皮酚的潜在靶点进行系统性挖掘(见图2)。借助SwissTargetPrediction与UniProt数据库验证[6],实验数据证实该成分具备与200余种人类蛋白靶点产生相互作用的潜力。利用STRING数据库进行的蛋白网络拓扑学分析表明,其核心作用节点覆盖核因子ĸB(nuclear factor-ĸB, NF-ĸB)、丝裂原活化蛋白激酶(mitogen-activated protein kinase , MAPK)、磷脂酰肌醇3-激酶(phosphatidylinositol 3-kinase , PI3K)/蛋白激酶B(protein kinase B ,AKT)等重要信号转导系统,这些枢纽网络与炎症级联反应、氧化损伤修复及细胞增殖调控存在密切关联,目前已完整绘制其多维度作用网络框架。
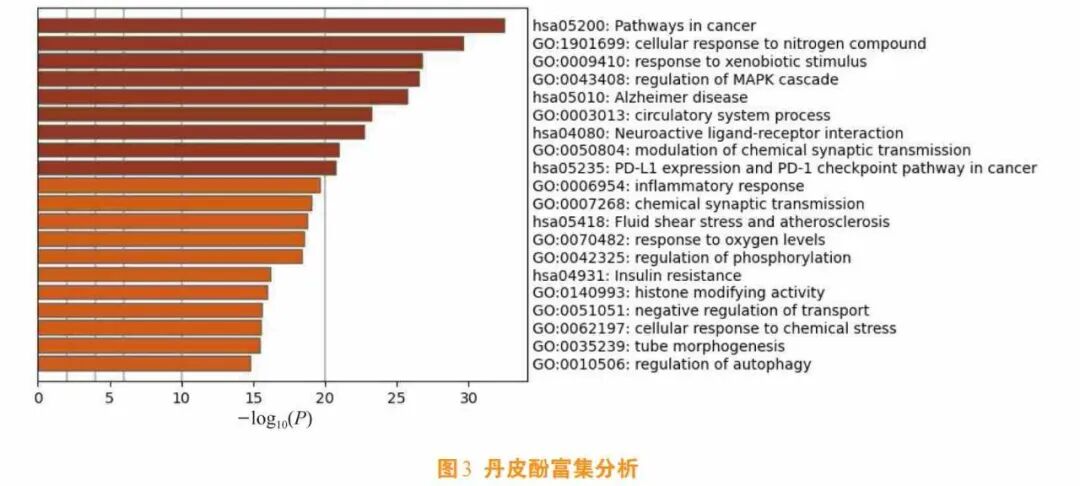
通路富集解析显示,丹皮酚作用靶点显著聚集于肿瘤发生通路(hsa05200)、炎症调控模块(GO:0006954)及神经退行病理进程(hsa05010),揭示其通过多靶点协同产生治疗效应的分子基础(见图3)。这种“微量成分-广谱调控”的特性,可解释其在抑制炎症风暴、阻断氧化应激瀑布效应及干预细胞周期紊乱等方面的整合调节能力,与中医“异病同治”理论形成分子层面的呼应——通过核心信号节点的网络化调控,实现对不同病理状态的协同干预。这种作用模式为阐释传统药效物质基础的现代科学内涵提供了创新视角。
2.1 抗炎作用
炎症[7]是一种由免疫细胞引起的复杂病理状况,其与很多疾病的发病机制相关。丹皮酚的抗炎活性已在多种疾病模型(包括动脉粥样硬化[8]、结肠炎[9]、神经炎症[10]等)中得到系统性验证。其作用机制涉及氧化应激调控、炎症信号通路干预及线粒体功能调节这3个层面。
研究表明[11],丹皮酚不仅能够通过提高超氧化物歧化酶(superoxide dismutase,SOD)活性、降低丙二醛(malondialdehyde,MDA)含量发挥抗氧化作用,还可以通过清除体内自由基如羟基自由基(·OH)、超氧阴离子自由基(O2-·)和2,2-二苯基-1-苦基肼自由基(2,2-diphenyl-1-picrylhydrazyl,DPPH)等,治疗过度氧化造成的机体损伤。在张鹏等[12]进行的DPPH自由基清除实验中,丹皮酚通过接受自由基中的单电子,使其配对并消除自由基,从而降低自由基对细胞的损伤,且其抗氧化能力呈剂量依赖性。
丹皮酚通过调节血脂、降低血液黏稠度及减轻肝脏脂肪沉积,激活腺苷一磷酸激活的蛋白激酶(adenosine monophosphate-activated protein kinase, AMPK)/AKT/哺乳动物雷帕霉素靶蛋白(mamma-lian target of rapamycin , mTOR)/自噬相关基因(autophagy-related gene ,ATG)通路并上调肉碱棕榈酰转移酶-1(carnitine palmitoyltransferase1, CPT 1)和沉默信息调节因子1(sirtuins 1 ,SIRT 1),抑制固醇调节元件结合蛋白-1c(sterol regulatory element binding protein-1c , SREBP-1c),从而缓解活性氧(reactive oxygen species ,ROS)介导的氧化应激[13]。其还可通过激活核因子E2相关因子2(nuclear factor erythroid 2-related factor 2 , Nrf2)/Kelch样ECH关联蛋白1(Kelch-like ECH- associated protein 1 , Keap1)/抗氧化反应元件(antioxidant response element ,ARE)信号通路增强细胞抗氧化能力[14],并直接减少ROS生成[15]。线粒体形态异常会导致线粒体功能障碍和ROS过量,从而导致线粒体膜电位的丧失,同时线粒体损伤也会促使ROS过度产生[16]。研究表明[17],丹皮酚可以抑制超氧化物产生、改善线粒体功能障碍,下调细胞凋亡相关蛋白的表达,缓解脂多糖(lipopolysaccharide , LPS)引起的肝细胞损伤。丹皮酚[18]还可通过过氧化物酶体增殖物激活受体γ共激活因子1α(peroxisome proliferator-activated receptor-γ coactivator-1α , PGC-1α)对线粒体依赖性凋亡产生显著影响。酪蛋白酶途径[19]将线粒体损伤与酪蛋白酶激活联系起来,并作为线粒体膜功能损伤的指标。研究发现[20],丹皮酚可以诱导线粒体膜去极化,并加剧了造血干细胞(hematopoietic stem cell ,HSC)中线粒体介导的细胞凋亡和激活酪蛋白酶级联反应,进而抑制HSC生长。
2.2 神经保护作用
丹皮酚在外周神经系统中发挥了重要的抗炎作用,其在许多中枢神经系统疾病中也发挥着显著作用。
糖尿病脑病(diabetic encephalopathy ,DE)作为糖尿病并发症中最常见的一种神经类疾病,其主要特征是认知功能受损,以及血糖水平升高引发的神经元损伤[21]。研究显示[22],丹皮酚可减轻氧葡萄糖剥夺(oxygen and glucose deprivation , OGD)所致的海马神经元的损伤。科学家在D-半乳糖诱导的脑老化模型中发现,丹皮酚可提升乙酰胆碱、谷胱甘肽(glutathione , GSH)及Na+-K+-ATP酶活性,改善DE小鼠的学习和记忆功能。脑源性神经营养因子(brain-derived neurotrophic factor , BDNF)和胰岛素样生长因子(insulin like growth factor ,IGF)水平的升高与神经营养系统认知障碍的病程进展风险呈负相关,二者可通过神经保护作用抑制认知功能衰退的进程。丹皮酚通过抑制大鼠海马体和皮质中酪蛋白酶-3的表达,并恢复BDNF和IGF的表达,从而增强认知功能保护[23]。丹皮酚也可以降低海马体的MDA和ROS水平,并下调糖尿病引起的阿尔茨海默病模型大鼠的促炎因子骨髓过氧化物酶(myeloperoxidase ,MPO)、TNF-α和IL-6表达,从而抑制星形细胞增殖[24]。
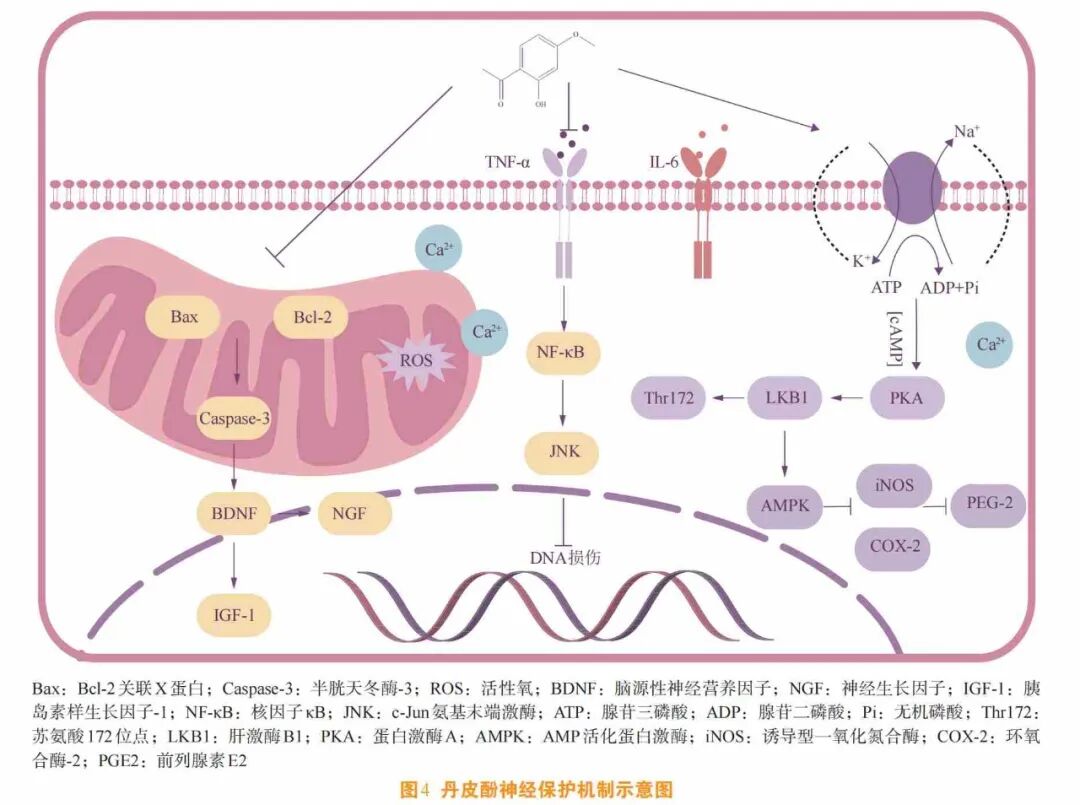
同时,丹皮酚可通过改善神经炎来保护中枢神经系统。在LPS诱导的小胶质细胞及小鼠巨噬细胞的慢性神经炎模型中[25],丹皮酚可通过阻断c-Jun氨基端激酶(c-Jun N-terminal kinase ,JNK)和NF-ĸB上游信号通路减少炎症因子TNF-α和IL-6的释放,降低一氧化氮合酶(nitric oxide synthase,NOS)和环氧合酶-2(cyclooxygenase-2 , COX-2)的水平,从而降低NO和前列腺素E2(prostaglandin E2 , PEG2)的表达,减轻神经炎。此外,丹皮酚一方面可通过激活肝激酶B1(liver kinase B1 , LKB1)和苏氨酸172(threonine 172, Thr172)的磷酸化来促进AMPK的激活,从而抑制LPS和干扰素γ(interferon-γ, IFN-γ)诱导的诱导型NOS(inducible NOS ,iNOS)、COX-2和血红素氧合酶-1(heme oxygenase-1,HO-1)的增加;另一方面,可抑制糖原合成酶激酶3α/β(glycogen synthase kinase-3α/β , GSK-3α/β)和ROS的产生[26](见图4)。
2.3 心血管保护作用
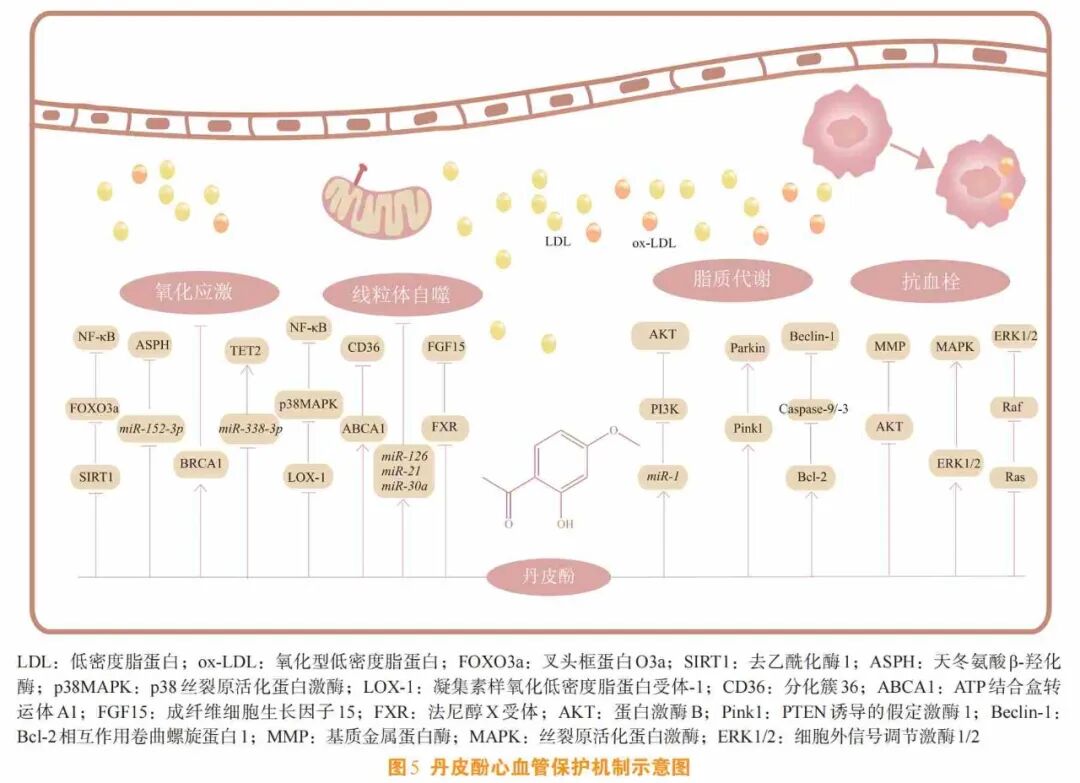
心血管疾病是全球范围内的主要死因之一。丹皮酚可通过以下几方面保护心血管:减少氧化应激、调节线粒体自噬、促进脂质代谢、抗血栓。
氧化应激由氧化-抗氧化失衡引发,导致ROS过度积累。心血管疾病中,低密度脂蛋白(low- density lipoprotein ,LDL)沉积于动脉内膜形成氧化型LDL(ox-LDL),刺激内皮细胞分泌炎性因子,诱导单核细胞黏附、迁移进入动脉内膜并转化为巨噬细胞,最终形成泡沫细胞,从而驱动动脉粥样硬化发展[27]。
Tang等[28]发现,丹皮酚通过调节SIRT1/叉头盒转录因子3a(FOXO3a)/NF-ĸB信号通路,在高葡萄糖/高棕榈酸处理的人脐静脉内皮细胞(human umbilical vein endothelial cell , HUVEC)中可提高细胞生存能力、SOD活性,并抑制细胞凋亡及ROS和炎症细胞因子的产生。其中,FOXO3a是一种在调节氧化应激方面发挥着重要作用的转录因子。Yu等[29]发现,丹皮酚可下调miR-338-3p并增加一种通过催化DNA5-甲基胞嘧啶的氧化发挥作用的酶TET2表达,从而抑制小鼠的LDL诱导的血管内皮细胞(vascular endothelial cell,VEC)凋亡、炎症和氧化损伤。天冬氨酸-β-羟化酶(aspartate- β-hydroxylase ,ASPH)作为一种聂型跨膜蛋白,不仅可以增强细胞增殖、迁移和入侵,还可通过刺激血管生成和免疫抑制来促进肿瘤生长。Liu等[30]发现,丹皮酚通过调节miR-152-3p/ASPH信号通路,抑制了由ox-LDL诱导的小鼠VEC凋亡,并降低了IL-1β、IL-6和ROS的表达水平。乳腺癌易感蛋白1(breast cancer susceptibility protein 1 ,BRCA1)基因作为基因组稳定性的调节器,参与DNA的修复和转录。Zheng等[31]发现,在心肌缺氧/再氧(myocardial anoxia/reoxygenation)损伤后,丹皮酚可通过上调BRCA1的表达来改善抗氧化酶的活性,并减少ROS的产生。
自噬作为细胞内关键的质控系统,通过清除受损细胞器维持心肌稳态。丹皮酚通过精细调控自噬维持心肌稳态[32]。适度的自噬可以缓解心肌损伤,在阿霉素诱导的心肌病模型中,丹皮酚通过上调miR-1抑制PI3K/AKT/mTOR通路,适度增强自噬活性以减少心肌细胞凋亡[33],同时激活Pink1/Parkin信号通路,促进模型大鼠的心肌线粒体自噬,减少缺氧引起的心肌细胞损伤[34]。然而,过度氧化应激会导致自噬关键启动因子Beclin-1过表达,细胞自噬被过度激活,促进细胞器的过度降解,导致细胞死亡。Beclin-1是一种Bcl-2相互作用蛋白,也是一种重要的自噬启动蛋白,可与其他自噬调节蛋白结合形成蛋白复合物,调控自噬水平。Tsai等[35]发现,丹皮酚通过上调Bcl-2蛋白,下调胱天蛋白酶-9/3及Beclin-1的表达,抑制自噬过度活化,改善心脏功能,从而改善心肌缺血再灌注损伤(myocardial ischemia-reperfusion injury) 引起的心律失常。
LDL氧化过程中产生的过氧化脂可能会直接损害内皮细胞。丹皮酚可阻断LOX-1/ROS/p38/MAPK/NF-κB信号通路,抑制ox-LDL诱导的内皮细胞凋亡[36]。此外,丹皮酚可通过上调miR-126、miR-30a表达以及下调miR-21表达[37],来抑制单核细胞对ox-LDL损伤的血管内皮细胞的黏附。ox-LDL通过诱导血管平滑肌细胞和巨噬细胞泡沫化,在动脉粥样硬化的发展中起着关键作用,而该疾病的特征正是血管壁中的脂质沉积和炎症。泡沫细胞的形成主要是由巨噬细胞的胆固醇外流受损或巨噬细胞对ox-LDL的不受控制的摄取引起的。巨噬细胞膜上有几种类型的清道夫受体(scavenger receptor,SR),其中A类SR(SR-A)和分化簇36(CD36)主要负责ox-LDL的吸收。然而,ATP结合盒转运体A1(ATP binding cassette transporterA1, ABCA1)、ATP结合盒转运体G1(ATP binding cassette transporter G1 , ABCG1)和B类SR(SR-B)主要负责细胞内脂质外流[38]。Li等[39]发现,丹皮酚通过促进ABCA1依赖性胆固醇排放和抑制CD36依赖性ox-LDL的吸收,减少了RAW264.7巨噬细胞中胆固醇的积累,并在载脂蛋白E基因敲除(ApoE_/_)小鼠中上调HO-1表达,增强主动脉ABCA1的表达、抑制CD36的表达,从而缓解动脉粥样硬化病变。
VSMC的异常增殖、迁移和表型转变是VSMC功能障碍的突出表现,可导致新内膜的形成。血小板衍生生长因子(platelet-derived growth factor, PDGF)是一种强效生长因子,可以促进有丝分裂。其中,PDGF-BB可促进体外培养的VSMC的增殖和迁移,激活细胞信号级联反应,从而促进细胞生长。研究发现,针对PDGF-BB诱导的VSMC增殖,丹皮酚能以剂量依赖性方式阻滞细胞周期G0/G1至S期进程[40]。Chen等[41]发现,丹皮酚可减少高葡萄糖对VEC的损害,通过抑制VEC释放血管内皮生长因子(vascular endothelial growth factor, VEGF)和PDGF,阻断VSMC内Ras/Raf/ERK1/2信号通路进而抑制VSMC的增殖(见图5)。
2.4 抗肿瘤作用
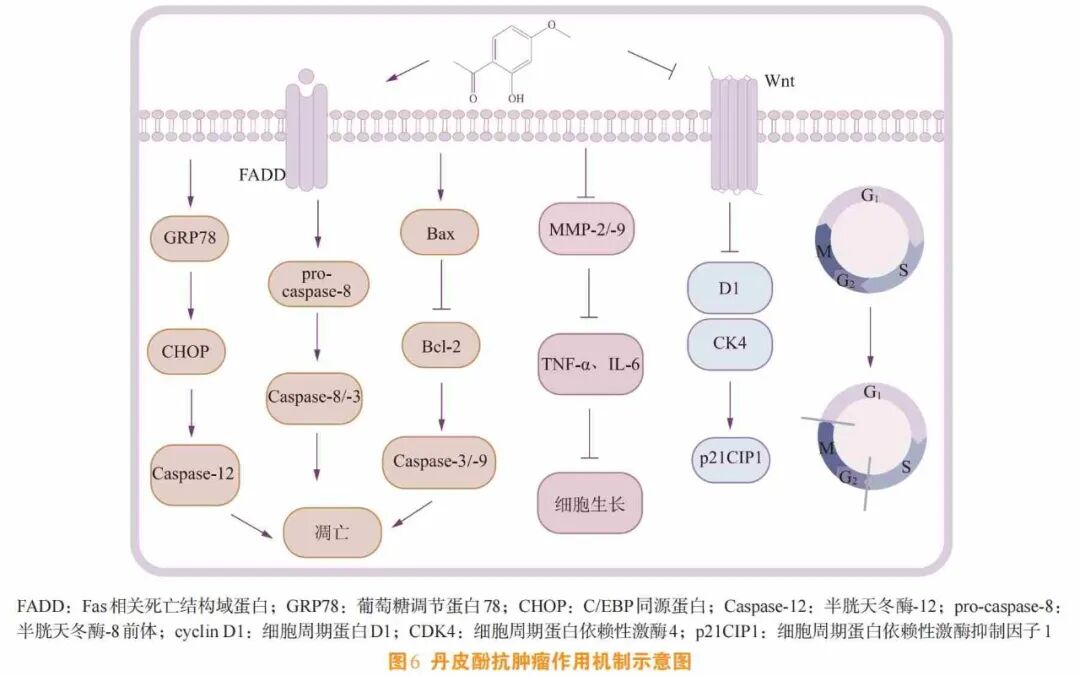
丹皮酚在抗肿瘤中的研究主要涉及结直肠癌、胃癌、肝癌、口腔癌等方面。研究人员发现[42],丹皮酚可通过抑制肿瘤细胞增殖、诱导细胞周期停止、诱导参与凋亡的基因表达、启动细胞凋亡发挥抗肿瘤作用。
细胞增殖[43]是生物体生长和发育的重要过程,同时也是许多疾病(如癌症)的基础。肿瘤的形成与细胞增殖存在着紧密的关系,肿瘤细胞具有不受控制的增殖能力,这是肿瘤生长和扩散的基础。因此,抑制肿瘤细胞增殖成为抗肿瘤治疗的关键策略之一。研究表明[44],丹皮酚可阻滞细胞周期进程、促进凋亡相关基因表达、激活细胞程序性死亡,下调促炎细胞因子(TNF-α、IL-1β、IL-6)分泌,从而抑制B16F10黑色素瘤转移,该机制与NF-κB通路的抗炎作用相似。同时,丹皮酚可下调基质金属蛋白酶-2/-9的蛋白表达,以浓度依赖的方式抑制人胃腺癌BGC823细胞的生长、迁移和入侵。
诱导肿瘤细胞发生周期阻滞是一种重要的抗肿瘤策略。许多外源性因素可引起细胞周期阻滞,从而发挥诱导细胞凋亡的作用。丹皮酚可诱导肿瘤细胞S期停滞进而引发凋亡,或阻断G0/G1期进程从而抑制肿瘤细胞的DNA复制及增殖[45]。Wnt/β-catenin信号通路[46]的异常激活可诱导参与细胞周期的胞核中的T细胞因子(T cell factor ,TCF)和淋巴样增强结合因子(lymphoid enhancer binding factor ,LEF)转录。研究人员进一步发现,丹皮酚通过抑制Wnt/β-catenin信号通路的激活下调细胞周期蛋白D1(cyclin D1)和细胞周期蛋白依赖性激酶4(cyclin-dependent kinase 4 ,CDK4),并上调细胞周期依赖性蛋白激酶抑制因子1A(cyclin- dependent kinase inhibitor 1A ,p21),从而将细胞周期阻滞于G0/G1期。同时,在结直肠癌研究中,丹皮酚能将HCT116细胞阻滞在G0/G1期,并导致S期和G2/M期细胞比例减少,这与在其他癌细胞系中的发现一致[47]。
作为肿瘤治疗的研究热点之一,诱导肿瘤细胞凋亡主要涉及的基因有Bcl-2基因、Bax基因、Fas基因、Caspase基因等,其中Bcl-2是凋亡抑制基因中的关键调控因子,可延长细胞寿命,促进细胞增殖,从而抑制细胞凋亡。Bax具有促凋亡作用,是Bcl-2基因家族成员之一,其编码的蛋白可与Bcl-2蛋白形成异源二聚体。当Bax过量表达时,可抑制Bcl-2的功能,从而促进细胞凋亡[48]。Caspase-3是Caspase家族中与细胞凋亡相关的重要成员。研究表明[49],丹皮酚可显著抑制Bcl-2表达,上调促凋亡蛋白Bax水平,使Bcl-2/Bax比值降低,同时激活Caspase-3、Caspase-8和Caspase-9以促进细胞凋亡。研究证明[50],丹皮酚通过促进细胞凋亡蛋白葡萄糖调节蛋白78(glucose-regulated protein 78, GRP78)、Caspase-3的表达及Ca2+的释放,引发线粒体功能障碍,进一步诱发细胞凋亡(见图6)。
丹皮酚作为中药牡丹皮的核心活性单体,其多靶点药理活性已得到广泛研究。本文对丹皮酚在不同种属中的吸收、分布、代谢和排泄(absorption, distribution, metabolism and excretion ,ADME)特征进行了总结,并探讨基于生理的药代动力学模型(physiologically based pharmacokinetic model ,PBPK model,PBPK模型)在预测中的应用前景。研究显示,大鼠体内丹皮酚的生物利用度在灌胃(28.92%)、肌肉注射(68.68%)、鼻腔给药(52.37%)间存在显著差异。丹皮酚溶解度较低,生物利用度差,在热水中溶解,可能会随着蒸汽蒸发。因此,丹皮酚需要在干燥和密封的环境中适当保存。由于其酚醛羟基的存在,丹皮酚具有较弱的酸性和一定的还原性,易氧化。首过效应明显、半衰期短、生物利用度低,是造成丹皮酚临床应用受限的主要原因。
3.1 吸收
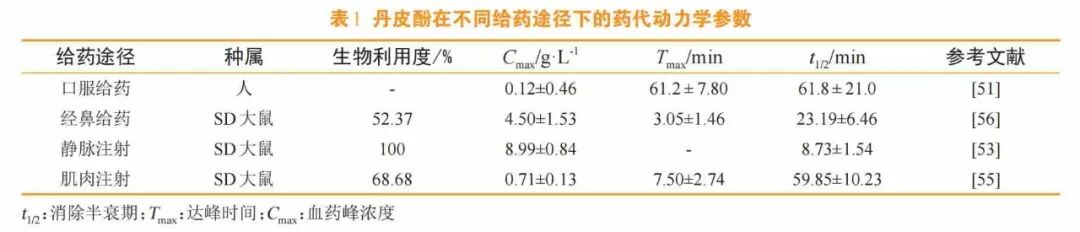
健康志愿者口服160mg丹皮酚后,消除半衰期(t1/2)和达峰时间(Tmax)均约为1h,血药峰浓度(Cmax)约为0.115mg·L-1(见表1)[51]。丹皮酚在肝微粒体中迅速代谢,30min的代谢残留含量仅为20.97%,丹皮酚在人工胃液和肠液中的t1/2分别是其在肝微粒体中的45倍和135倍[52],这表明丹皮酚虽在胃肠道中稳定,但进入肝脏后会被迅速代谢,导致生物利用度降低,需通过优化剂型或调整给药途径来规避肝脏首过代谢。给大鼠静脉注射4mg·kg-1的丹皮酚[53],其吸收迅速,代谢消除较快,半衰期较短,血药浓度-时间过程符合二房室模型。孙言才等[54]发现,对小鼠灌胃给药50mg·kg-1的丹皮酚7min后,在血浆中可检测到丹皮酚及其代谢产物,丹皮酚Tmax为(6.30±0.70)min,t1/2为(41.49±2.96)min。基于丹皮酚口服生物利用度低的问题,研究人员探索了其他给药途径:例如,在给大鼠肌肉注射丹皮酚(10mg·kg-1)后,通过高性能液相色谱测量血浆中丹皮酚的浓度,其t1/2为(59.85±10.23)min,Tmax为(7.50±2.74)min,给药后丹皮酚的生物利用度为68.68%[55],但注射中使用的高浓度吐温80(Tween-80)和乙醇可对肌肉和血管产生强烈的刺激;另有研究发现,鼻腔内给药后,丹皮酚生物利用度可达到52.37%,高于灌胃给药的生物利用度。这些研究为优化丹皮酚的给药途径提供了新的方向。
3.2 分布
丹皮酚进入体内后迅速分布,其在体内的分布特点尚未得到全面阐述,但已有研究表明,丹皮酚可能通过血液循环到达全身各组织器官,发挥其药理作用。对小鼠灌胃给药65mg·kg-1的丹皮酚后,其在体内主要分布于肝中,其次是脾、小肠上端、肾、结肠、心、肺等组织[57]。利用HPLC-UV法测定大鼠不同组织中的丹皮酚浓度,发现灌胃给药194.4mg·kg-1的丹皮酚后,其在肝、肾中含量较高,在心、脾、肺和大脑均可检测到丹皮酚[58]。此外,有研究表明[59],丹皮酚可通过血脑屏障,并在大脑的某些区域具有药理活性。以上研究为丹皮酚对肝脏的保护作用、在脾脏中的免疫调节作用以及对心脏和大脑氧化损伤的预防潜力提供了依据。丹皮酚的组织分布具有显著器官选择性。新型检测技术与靶向递送系统的结合,有望突破丹皮酚的分布限制,推动其临床转化。
3.3 代谢与排泄
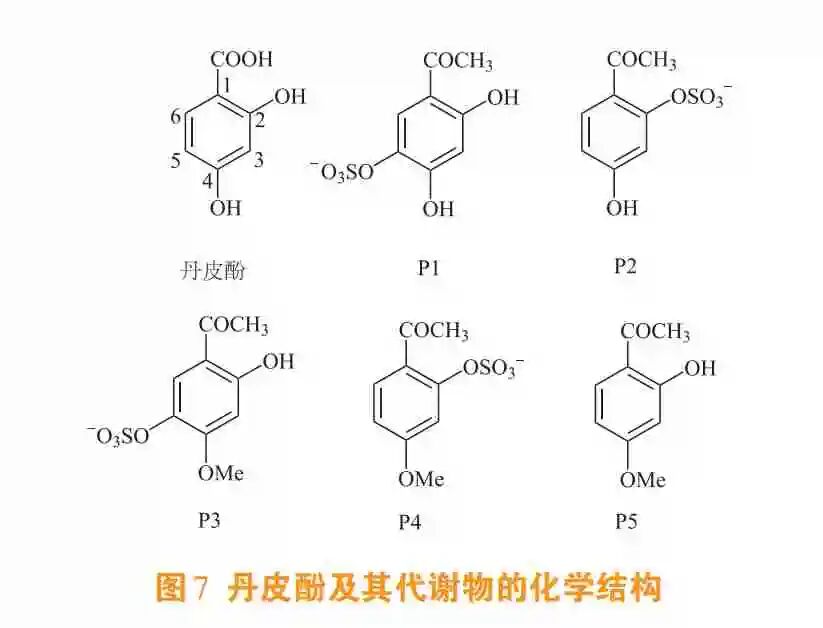
丹皮酚在给大鼠经口使用后会迅速吸收和排泄。Yasuda等[60]在给大鼠经口使用丹皮酚后,首次在其尿液中鉴定出丹皮酚的5种代谢产物,即2,4-二羟基苯乙酮-5-O-硫酸酯(P1)、重乙酰苯酮-2-O-硫酸酯(P2)、2-羟基-4-甲氧基苯乙酮5-O-硫酸酯(P3)、丹皮酚-2-O-硫酸酯(P4)和重乙酰苯甲酮(P5)(见图7)。
研究表明,在丹皮酚灌胃5min后的大鼠的血浆中,可识别出4种代谢物P2~P5,且这些代谢物在灌胃后约20min达到最大血浆浓度,这表明丹皮酚可迅速代谢成P2~P5等代谢物。因此丹皮酚的口服生物利用度差可能是由于其在血液中存在首次代谢[61]。丹皮酚在体内的代谢消除也较快,其代谢途径包括被羟基化为2,5-二羟基-4-甲氧基乙酰苯酮并脱甲基化,以及被羟基化后再进行硫酸化这两种。胡欣彤[62]等研究发现,对大鼠灌胃给予牡丹皮提取物后,丹皮酚的代谢产物主要经尿液、胆汁和粪便排泄,且排泄量依次递减。
3.4 基于生理的药代动力学模型在丹皮酚研究中的应用潜力
PBPK模型是一种基于生理学参数动态模拟药物在体内的吸收、分布、代谢和排泄过程的工具,目前已广泛应用于药物生物等效性研究[63]。
利用PBPK模型,可预测丹皮酚在不同给药途径下的生物利用度,用于指导纳米载体或脂质体开发[64]。同时,可整合丹皮酚在不同种属动物(如大鼠、犬)中的实验数据来预测人体药代动力学参数,评估种属间代谢差异对药效/毒性的影响,减少临床试验剂量探索成本[65]。丹皮酚具有多组织分布的药代动力学特征。基于此特征,可构建模型以预测其在血浆和脑组织的药物浓度,并评估其在心血管系统等特定组织中的潜在蓄积情况。此外,在药物相互作用与个体化用药方面,PBPK模型可用于预测丹皮酚与CYP450抑制剂/诱导剂联用时的血药浓度变化。如今,人工智能技术已被广泛应用于药物靶点识别、ADME预测及临床试验设计等药学领域[66],并对PBPK建模产生深远影响。因此,结合机器学习优化PBPK模型参数,有望为丹皮酚的深入研究带来新的突破。
本文全面探讨了丹皮酚的核心药理活性及其作用机制。研究显示丹皮酚具有多维度生物活性,涵盖炎症调节、神经功能维护、心血管系统修复及肿瘤抑制等方面。然而,尽管丹皮酚各种药理作用均涉及抗氧化与抗炎等共性机制,但其在不同病理条件下的主导作用通路呈现明显特异性。
在炎症调控方面,丹皮酚可显著抑制ROS生成及炎症介质释放,在急性炎症模型的治疗中展现快速起效特点;在神经系统保护方面,丹皮酚可改善线粒体功能和促进神经营养因子合成,对慢性神经退行性病变有干预优势;在心血管保护方面,丹皮酚可修复血管内皮与调节脂质代谢,表现出组织特异性的保护作用;丹皮酚的抗肿瘤活性主要源于细胞周期阻滞和Caspase依赖性凋亡通路的激活。同时,丹皮酚“清热凉血、活血散瘀”的传统功效与现代分子机制存在显著关联性,其清热作用对应NF-ĸB介导的炎症因子抑制机制,而活血功能则通过血管内皮功能改善和脂代谢调节实现,这与现代医学对血瘀证病理特征的认识高度契合。
药代动力学研究揭示了给药途径对丹皮酚生物利用度的显著影响,这为其新型递药系统的开发提供了理论支撑。临床应用显示丹皮酚对发热性头痛、神经痛等急症及湿疹等皮肤病症具有明确疗效,但受限于其水溶性差、口服生物利用度低、稳定性差和室温下挥发性高等缺陷,丹皮酚除了抗炎活性(以软膏形式应用于皮肤炎症治疗)外,其他药理活性尚未得到临床应用。采用纳米脂质体包封、环糊精包合及经鼻给药等新型制剂技术可有效改善丹皮酚的稳定性和靶向性。
基于网络药理学分析,丹皮酚对“热毒血瘀”症候相关疾病有精准治疗的潜力。丹皮酚现有研究多集中于动物实验阶段,临床数据仍较匮乏,毒理学评价及长期用药安全性仍需深入探讨,未来研究或将聚焦代谢产物活性分析、种属差异性研究,以及智能化药代动力学模型在个体化给药中的应用探索。
本文引文格式:
赵紫雅, 孙建国. 丹皮酚的药理作用及药代动力学研究进展[J]. 药学进展, 2026, 50(3): 254-263.
信息来源:药学进展




免责声明
“汇聚南药”公众号所转载文章来源于其他公众号平台,主要目的在于分享行业相关知识,传递当前最新资讯。图片、文章版权均属于原作者所有,如有侵权,请在留言栏及时告知,我们会在24小时内删除相关信息。
本平台不对转载文章的观点负责,文章所包含内容的准确性、可靠性或完整性提供任何明示暗示的保证。
喜欢的点个“看一看”和"喜欢"吧

不然微信推送规则改变,有可能每天都会错过我们哦~





