 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库PNAS:天津医科大学张锴团队发现ENO1入核耦合HDAC1通过局部PEP抑制驱动组蛋白乳酰化
组蛋白去乙酰化酶(HDACs)在转录调控中发挥重要作用,并主要在经典转录复合体内催化去乙酰化反应。然而,其他HDAC相关蛋白在调控这一过程中的机制作用仍未完全阐明。
2026年6月17日,天津医科大学张锴独立通讯在PNAS在线发表题为
“ENO1 couples HDAC1 to regulate histone lactylation and gene transcription”
的研究论文。该研究为系统解析活细胞中内源性HDAC互作网络,作者开发了BimPL,这是一种由双功能分子介导的邻近标记策略。
利用BimPL结合定量蛋白质组学,作者不仅稳定捕获了已知的HDAC复合体,还鉴定出潜在的新互作蛋白,其中包括糖酵解酶烯醇化酶1(ENO1)。重要的是,作者发现ENO1能够转位进入细胞核,并在染色质水平与HDAC1发生相互作用;而这一过程通过局部生成的磷酸烯醇式丙酮酸(PEP)抑制HDAC1活性。由此,ENO1–HDAC1的耦合促进组蛋白赖氨酸乳酰化(Kla)的增加,从而驱动肝脏
恶性肿瘤
中致癌基因的转录重编程。本研究建立了BimPL作为解析内源性蛋白互作组的有效工具,并揭示了一种由代谢酶调控HDAC活性的机制,从而调控组蛋白乳酰化修饰,强调了ENO1作为“多功能(月光型)蛋白”在表观遗传重编程中的关键作用。

哺乳动物组蛋白去乙酰化酶(histone deacetylases, HDACs)是一类关键的转录调控因子,可分为两个家族:NAD依赖性的sirtuins(III类)以及金属依赖性的HDACs,其中包括I类(HDAC1、2、3、8)、II类(HDAC4、5、6、7、9、10)以及IV类(HDAC11)。HDACs通常通过催化去除底物蛋白赖氨酸残基上的乙酰基或其他酰基修饰来调控基因转录,其功能异常与多种人类疾病相关,包括癌症。HDACs的功能很大程度上取决于其动态组装形成不同的表观遗传复合体。在这些复合体中,HDACs在DNA结合转录因子及其他伴侣蛋白的协同作用下,被时空特异性地招募到特定基因组位点,从而调控基因表达程序。因此,在天然状态下系统性鉴定HDAC相关蛋白对于理解其分子机制至关重要。
为解析HDAC复合体组成,已有多种方法被开发,包括基于肽探针、标签化HDAC以及固定化HDAC抑制剂等策略,用于捕获HDAC及其相互作用蛋白。例如,Dose及其同事利用将HDAC抑制剂基团连接在肽骨架上的探针,研究了内源性HDAC复合体的底物选择性与组成。然而,由于探针与HDAC复合体之间的相互作用通常是瞬时且非共价的,这些方法仍面临显著挑战。为解决这一问题,研究者进一步发展了光亲和标记策略,可将非共价相互作用转化为共价键,从而增强HDAC互作蛋白的捕获能力。但此类基于HDAC抑制剂的光探针主要适用于直接结合蛋白的标记,对HDAC复合体中的间接互作蛋白识别能力有限。近期还发展了一种DNA基础的亲和标记方法,通过设计不同长度DNA链的捕获探针,实现对间接HDAC相关蛋白的标记,但该方法主要应用于裂解样本,在细胞裂解过程中HDAC复合体可能发生解离,难以反映天然状态。
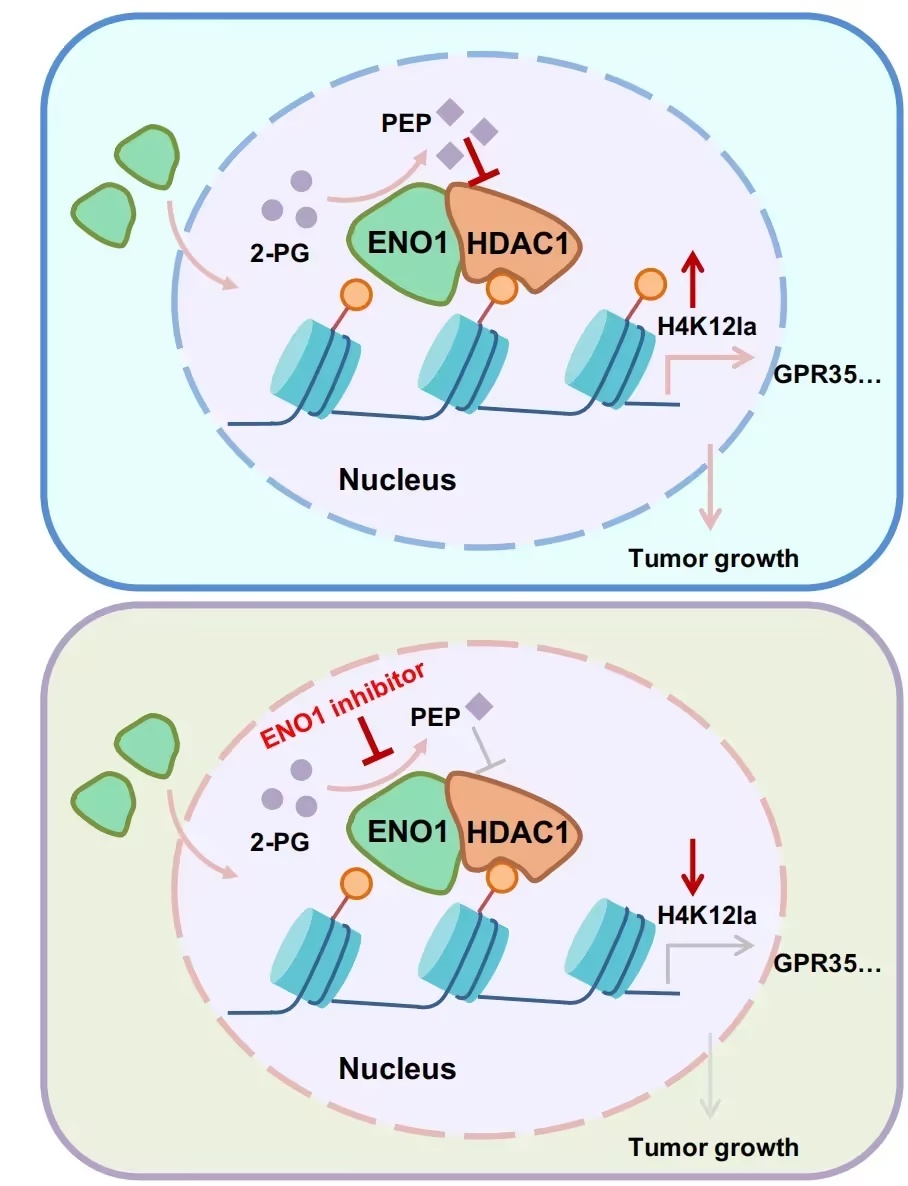
图1.全文总结图(摘自
PNAS
)
近邻标记(proximity labeling, PL)技术作为一种由工程化酶催化的策略,已成为在活细胞中捕获蛋白直接及间接互作的有力工具。该技术通常通过遗传编码方式,将APEX2或BioID等近邻标记酶与目标蛋白融合表达。然而,这些方法需要外源表达融合蛋白,可能扰乱天然蛋白复合体结构与功能,并带来异常定位及互作伪影风险。
为实现内源性蛋白复合体的天然状态解析,作者在此开发了BimPL,一种结合APEX2标记与HDAC靶向识别的化学生物学策略。该方法利用一种双功能分子探针,同时结合APEX2-SNAP与HDACs,从而在活细胞中实现空间限制性的互作蛋白生物素化标记。结合质谱分析,BimPL不仅可以稳定鉴定已知HDAC及其复合体,还发现了多个未被充分表征的互作蛋白,其中包括烯醇化酶1(ENO1)。
进一步研究表明,ENO1能够与HDAC1结合并发生共定位,通过局部生成磷酸烯醇式丙酮酸(PEP)抑制HDAC1活性,从而在特定位点驱动组蛋白乳酰化修饰的发生。值得注意的是,ENO1–HDAC1复合体更倾向于促进组蛋白H4K12乳酰化(H4K12la),进而促进基因表达与肿瘤发生。
本研究为在活细胞中解析内源性HDAC相关蛋白提供了一种新方法,同时揭示了核内ENO1与HDAC1通过调控组蛋白乳酰化(Kla)与基因转录的机制,从而拓展了对HDAC活性调控及代谢—表观遗传耦合机制的理解。
参考消息:https://doi.org/10.1073/pnas.2535245123





