 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库PNAS:突触释放的“双速变速箱”,西安交通大学刘浩文等团队揭示SNT-1和SNT-3如何用不同的分子接口分别控制快慢挡
在秀丽隐杆线虫(Caenorhabditis elegans)神经肌肉接点处,神经递质释放由一个双重Ca²⁺传感器系统调控,该系统由SNT-1和SNT-3组成,其功能类似于哺乳动物某些神经元中的Ca²⁺传感器系统,例如海马体中的突触结合蛋白1和7(Syt1/Syt7)。
2026年6月18日,西安交通大学刘浩文,香港城市大学胡志涛和伊利诺伊大学Janet E. Richmond共同通讯在
PNAS
在线发表题为
Evolutionarily conserved and divergent mechanisms of dual Ca2+ sensors in synaptic vesicle exocytosis
的研究论文。
该研究探讨了SNT-1和SNT-3如何通过其与SNARE复合体的潜在相互作用以及其多碱性基序(polybasic motifs)介导快速与缓慢的神经递质释放。基于AlphaFold 3对SNT-1–SNARE和SNT-3–SNARE复合体的结构预测显示,其C2B–SNARE构型与经典的Syt1–SNARE一级界面一致(Zhou et al., Nature 525, 62–67, 2015),并精确识别了C2B结构域以及SNAP-25和Syntaxin中的保守结合残基,提示该相互作用在进化上的保守性。
通过定点突变的电生理分析表明,SNT-1和SNT-3都依赖C2B–SNARE相互作用以及其C2结构域内的多碱性基序来驱动诱发型快速和缓慢神经递质释放。
值得注意的是,SNT-1和SNT-3在对C2B–SNARE界面不同区域以及各自多碱性基序的依赖性上存在差异,提示Ca²⁺触发的快速与缓慢释放通过不同的分子机制实现。此外,作者发现SNT-1通过多种途径介导自发性神经递质释放,不仅依赖于主要的C2B–SNARE界面,还涉及其他潜在的SNARE结合相互作用。
综上,本研究揭示了在秀丽隐杆线虫中由双Ca²⁺传感器调控的突触胞吐过程所具有的既保守又分化的分子机制。

神经递质通过突触小泡(synaptic vesicle, SV)胞吐从突触前神经末梢释放,该过程由SNARE(可溶性N-乙基马来酰亚胺敏感因子附着蛋白受体)复合体调控,而SNARE复合体是膜融合机器的核心组成部分。这一过程受到SNARE结合蛋白的精细调控,例如Munc13、Munc18以及突触结合蛋白(synaptotagmin)。
其中,Syt1作为一种突触小泡蛋白,已被确立为负责快速、同步神经递质释放的主要Ca²⁺传感器。其他突触结合蛋白基因,如Syt2和Syt9,也在特定神经元群中发挥快速Ca²⁺传感器的作用。
这些蛋白的典型结构包括:短的N端腔内区、跨膜结构域、无序连接区,以及串联的C2结构域——C2A和C2B,两者均通过其Ca²⁺结合环中保守的天冬氨酸残基结合Ca²⁺。近期研究鉴定Syt7为介导海马体中慢速、非同步释放的次级Ca²⁺传感器。
与Syt1不同,Syt7敲除不会影响基础SV释放,但会消除Syt1缺失神经元中残余的非同步释放。此外,在Syt1/Syt7双敲除神经元中,SV启动(priming)几乎完全丧失,而单独敲除任一基因时该过程仍然保留,提示Syt1与Syt7在启动过程中具有部分冗余功能。
总体而言,这些发现支持一种“双Ca²⁺传感器模型”:Syt1与Syt7分别作为快速与慢速Ca²⁺传感器,调控海马体SV释放的不同时间动力学成分。
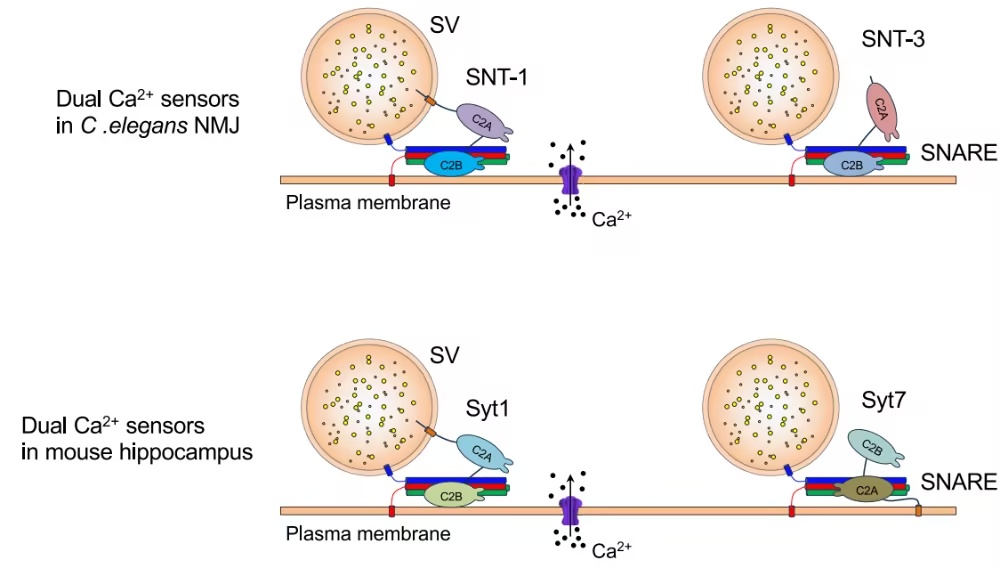
图1.蠕虫双Ca2+传感器的示意图模型NMJ (SNT-1和SNT-3)和小鼠海马(Syt1和Syt7)(摘自
PNAS
)
尽管已有二十年的研究,快速与慢速Ca²⁺传感器(例如海马体中的Syt1与Syt7)如何在分子层面精确调控SV胞吐的机制仍未完全阐明。生化与功能研究表明,Syt1的C2结构域可与SNARE复合体相互作用,这是SV融合过程中的关键步骤。
尽管Syt1 C2结构域及SNARE复合体的独立结构已被解析,但完整Syt1–SNARE复合体的精细结构模型长期缺失。近期Brunger团队首次解析了Syt1–SNARE复合体的晶体结构,揭示了Syt1 C2结构域与SNARE蛋白之间的多个接触界面。此外,位于Syt1–SNARE主要界面的关键残基突变会显著削弱Syt1功能。
Syt1 C2结构域与质膜的相互作用同样在触发SV释放中起关键作用。C2结构域可响应胞质Ca²⁺浓度升高,促使其Ca²⁺结合环的侧链快速插入富含负电荷磷脂的膜结构。已有研究表明,这种膜插入作用能够加速SNARE驱动的SV融合过程,无论是在体外体系还是培养神经元中均如此。
除了Ca²⁺结合环之外,两个C2结构域中的多碱性基序(polybasic motifs)也被证明能够与质膜相互作用。每个结构域中的四个连续赖氨酸残基形成多赖氨酸区域,可在Ca²⁺非依赖条件下与膜上的PIP2脂质簇结合。目前普遍认为,Syt1中的SNARE相互作用与膜相互作用在胞吐前同时发生,而Ca²⁺内流则促进这些相互作用并重塑膜结构,从而推动SV融合。
作者此前研究发现,在秀丽隐杆线虫(Caenorhabditis elegans)中存在一个双Ca²⁺传感器系统,由SNT-1(Syt1的同源蛋白)和SNT-3组成,共同触发诱发型快速与慢速SV释放。与线虫及哺乳动物中多数其他synaptotagmin不同,SNT-3独特地缺乏跨膜结构域(TMD)。
值得注意的是,TMD对于SNT-1作为快速Ca²⁺传感器的功能并非必需:在snt-1突变体中观察到的诱发EPSC延迟增加,可被缺失TMD的SNT-1完全恢复。C. elegans中的SNT-1/SNT-3系统在功能上与海马体Syt1/Syt7系统相似:缺失主要传感器会严重损害快速释放,而单独缺失次级传感器不会影响基础传递,但在主要传感器缺失时会消除残余释放。
此外,主/次Ca²⁺传感器在SV启动过程中具有冗余贡献。这些共同特征使秀丽隐杆线虫成为解析双Ca²⁺传感器介导突触传递进化保守机制的理想模型。
尽管已有上述研究,双Ca²⁺传感器触发SV胞吐的精确分子机制仍不清楚。在本研究中,作者分析了SNT-1和SNT-3中与Syt1–SNARE复合体结构中SNARE相互作用界面对应的保守残基。
作者的结果表明,该结构中鉴定的C2B–SNARE主要界面对SNT-1和SNT-3的功能均至关重要,但该界面内部不同亚区域对两种传感器的贡献存在差异。此外,它们C2结构域中的多碱性基序也发挥关键作用,但不同C2结构域之间的贡献方式不同。作者还发现,SNT-1参与多种潜在SNARE相互作用模式,共同支持自发性SV释放。
总体而言,本研究表明C. elegans中的双Ca²⁺传感器在触发SV胞吐时同时利用保守与分化的分子机制,为理解突触传递机制的进化提供了重要线索。
参考消息:https://doi.org/10.1073/pnas.2532992123





