 五度妙笔
五度妙笔 API商城
API商城
 数据库
数据库诺奖团队最新Science:AI设计“迷你”基因编辑酶,超越自然进化,高效编辑人类基因组


撰文丨王聪
编辑丨王多鱼
排版丨水成文
CRISPR-Cas 系统通过实现可编程的、序列特异性的 DNA 和 RNA 靶向,已彻底改变了基因组编辑技术,为精确的遗传和表观遗传调控奠定了基础。蛋白质设计有望进一步拓展这些功能,创造出具有自然界中未见的功能和特性(包括序列与结构)的 RNA 引导的核酸酶。
然而,这类设计极具挑战性,因为这些核酸酶是多结构域的核酸结合蛋白,其活性依赖于不同构象状态对 RNA 和 DNA 的协同识别、激活及切割。基于序列的生物语言模型通过进化数据训练,能够通过推断序列与功能之间的关系来生成 RNA 引导的核酸酶,但即使经过大量后续筛选,其所生成的活性核酸酶通常仍与训练时参考的序列高度相似。
结构指导的理性设计方法则提供了一种稳健策略,可采样出高度多样化的蛋白质序列,甚至包括自然界中不存在的结构,从而实现蛋白质的从头设计。尽管该方法在动态开关和 DNA 结合蛋白的设计上已取得成功,但对于具有多个功能结构域和构象状态的复杂酶(例如 RNA 引导的核酸酶)的设计,依然是一个极具吸引力却尚未解决的难题。
2026 年 7 月 15 日,CRISPR 基因编辑先驱、诺奖得主 Jennifer Doudna 教授在国际顶尖学术期刊 Science 上发表了题为:Structure and evolution-guided design of minimal RNA-guided nucleases 的研究论文。
该研究采用一种蛋白质设计策略,将结构导向的逆折叠模型与基于进化的氨基酸残基约束相结合,生成了活性强且序列多样的 TnpB 变体(最小的 CRISPR-Cas12 样核酸酶),命名为——SynTnpB。通过高通量筛选 AI 生成的变体,获得了在细菌、植物和人类细胞中保持甚至超过野生型活性的基因编辑工具。对其中序列差异化最大的变体进行冷冻电镜结构解析,揭示了其在不同构象下 RNA-DNA 界面中的稳定相互作用,证明了该方法的设计潜力。这项研究建立了一种创造非天然 RNA 引导的核酸酶及构象活性核酸结合蛋白的策略,显著扩展了可设计的蛋白质空间。

“迷你版”基因编辑工具
说到基因编辑,很多人首先想到的通常是被誉为“基因魔剪”的革命性工具——CRISPR-Cas9,但他的缺点也很明显——Cas9 核酸酶太大了,以至于难以通过腺相关病毒(AAV)进行递送,这大大限制了其在体内基因治疗中的应用。
于是,科学家们把目光投向了 TnpB——这是一类小得多的微型核酸酶,只有约 400 个氨基酸,不到 Cas9 一半大小。它是 CRISPR-Cas12 家族的祖先,虽然体型迷你,却能完成 RNA 引导的 DNA 切割任务。
但问题来了:自然界的野生型 TnpB 虽然能用,但还不好用。
AI 设计:不只是模仿,更是创造
传统上,想要改进这类蛋白质,科学家通常采用两种方法——
一种是基于序列的生物语言模型,它们能生成蛋白质序列,但往往只是对天然序列的“微调”,缺乏真正的创新。
另一种是结构指导的理性设计,可以创造出高度新颖的序列,但对于像 TnpB 这样需要协调多个功能域和构象状态的复杂酶来说,难度极大。
这次,研究团队玩了个“组合拳”。他们使用了名为 ESM-IF1 的反向折叠模型——这是一种 AI 模型,能够根据给定的蛋白质三维结构,逆向推导出可能折叠成该结构的氨基酸序列。
但光靠结构不够。研究团队还引入了进化信息约束:通过分析数千种天然 TnpB 的同源序列,找出哪些位点在进化中高度保守(说明很重要),哪些位点与 RNA 或 DNA 存在共进化关系(说明有相互作用)。他们将这两类关键位点“锁定”,其余位置则交给 AI 自由发挥。
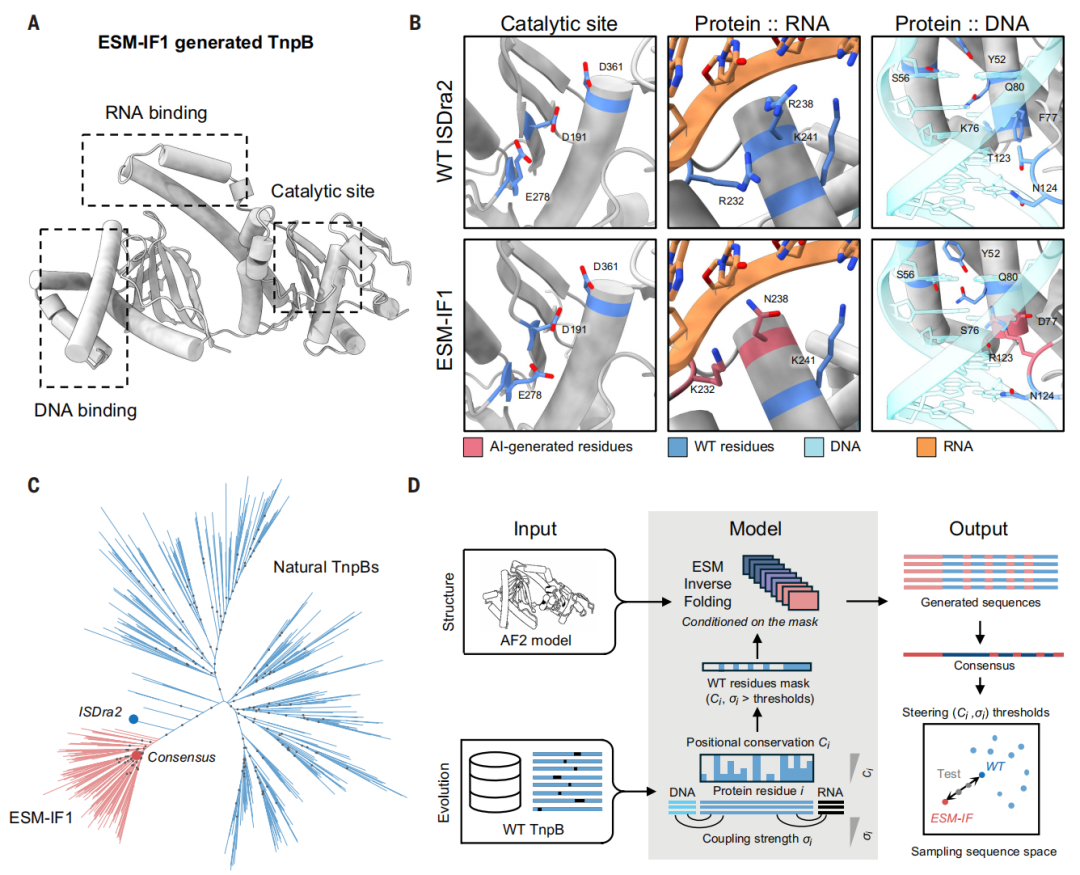
基于 ESM-IF1 的 RNA 引导的核酸酶 TnpB 设计策略
研究团队成功设计出了 466 个有活性的 TnpB 变体,成功率高达 24%,其中约 8% 的活性甚至超过了天然版本。
分域设计:分而治之的智慧
该研究中的一个有趣的发现是,TnpB 的不同结构域对序列变化的容忍度截然不同。
研究团队采用了“分域设计”策略:将 TnpB 分为 REC(识别结构域)和 NUC(核酸酶结构域)两个独立模块,分别由 AI 设计优化后,再进行组合。
结果显示,NUC 结构域非常“皮实”,即使在高达 0.65 的保守性阈值下,16 个变体中仍有 13 个保持活性。而 REC 结构域则相当“娇气”,只有在最低的保守性阈值(0.25)下才有一个变体保持活性。
这种不对称性反映了不同结构域的功能差异:REC 负责识别 DNA 上的 TAM 序列,需要精确的分子识别;而 NUC 主要负责切割,对序列变化的容忍度更高。
这一发现为未来设计其他多结构域蛋白提供了重要启示:对不同模块采取不同的设计策略,可能是成功的关键。
在人类细胞中的“超常发挥”
最激动人心的结果来自人体细胞实验。
研究团队选择了 9 个最具代表性的 AI 设计变体——v1-v9,在人类细胞系 HEK293T 中测试它们对蓝色荧光蛋白(BFP)基因的敲除效率。
结果显示,来自耐辐射奇球菌的野生型 ISDra2-TnpB 的编辑效率约为28%,而 AI 设计的 v1 和 v5 变体分别达到了 46% 和 50%——提升了近一倍!更令人振奋的是,在对内源性基因(RUNX1、EMX1等)的编辑中,v1 变体在 EMX1 位点的编辑效率是野生型的 3.8 倍,v5 则是 3.1 倍。在拟南芥原生质体中,v1 变体的的编辑效率同样由于野生型。此外,这些 AI 设计的变体保持了良好的特异性,没有出现大量脱靶效应。
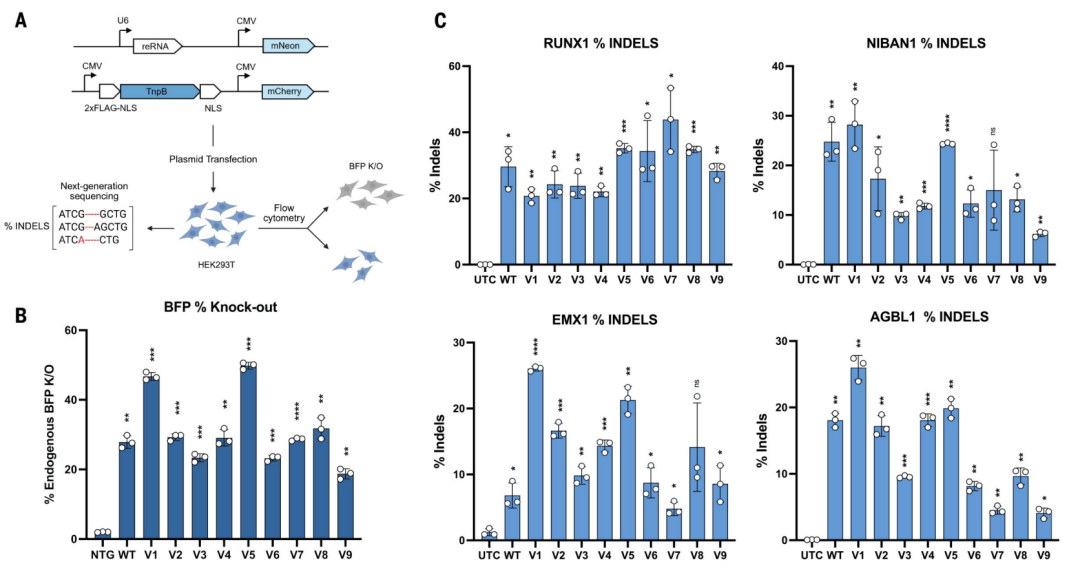
AI 设计的 TnpB 用于 HEK293T 细胞的基因编辑
这些 AI 设计的“基因剪刀”,在人类细胞以及植物细胞中的某些靶点上的编辑效果,大幅超越了自然进化了亿万年的野生型。
冷冻电镜下的“解剖学”
为了搞清楚 AI 到底做了什么,研究团队对活性最高且序列差异最大的 v7 变体进行了冷冻电镜结构解析。v7 变体与野生型仅有 77% 的序列同源性——也就是说,超过五分之一的氨基酸都被 AI 替换掉了。
冷冻电镜图像显示,在 RNA-蛋白质界面上,AI 引入了多个带正电荷的氨基酸替换(K224R、Q227R、T228R、K245R),形成了更强的静电相互作用网络,像是一双更有力的手紧紧握住 RNA 分子。
更令人惊讶的是,研究团队捕获到了一个此前从未在 TnpB 中观察到的转座子相关基序(transposon-associated motif,TAM,相当于 Cas9 的 PAM 序列)结合中间态。在这个状态下,蛋白质和 RNA 的构象类似于二元复合物,但已经与目标 DNA 有了初步接触。AI 设计的氨基酸残基稳定了这个短暂的过渡状态,使其得以被捕捉和研究。
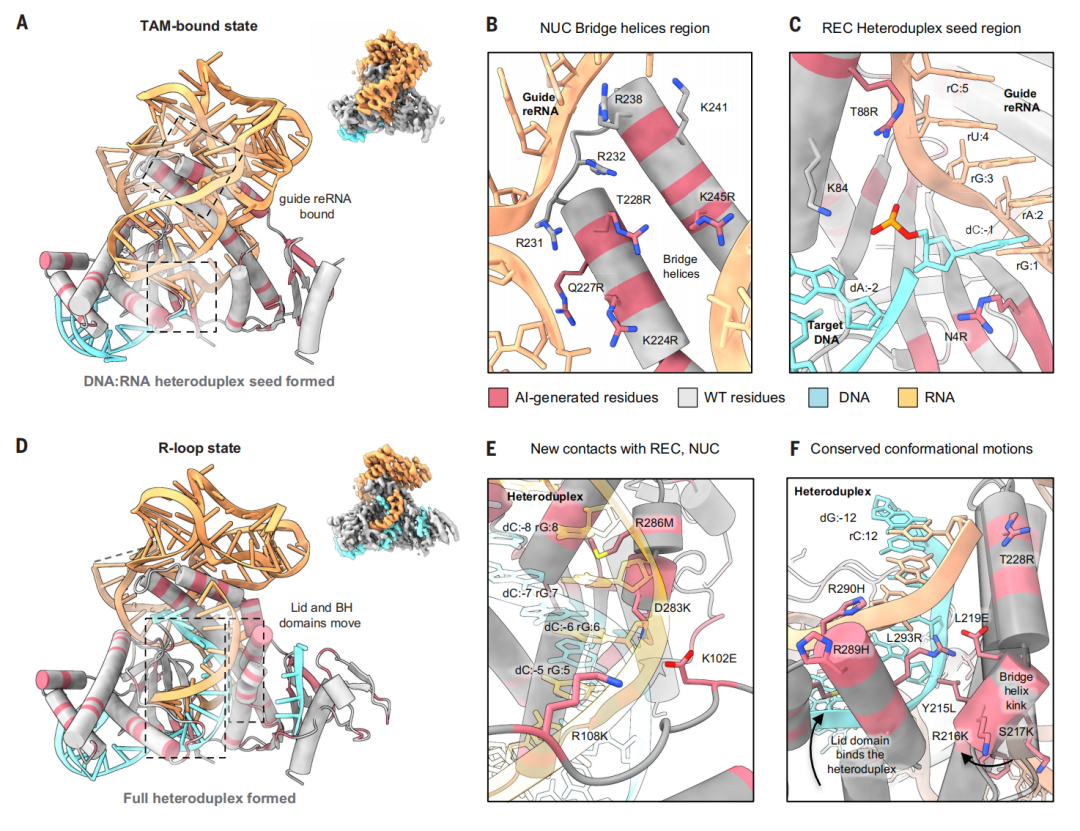
AI 设计的 v7 变体的冷冻电镜结构揭示了 RNA-DNA 界面残基及保守的构象运动
这表明,AI 不仅仅是在复制大自然的设计方案,而是在创造新的、更优的分子相互作用网络,甚至可能改变了蛋白质的动力学行为。
意义与展望
这项研究的突破性在于——
第一,它证明了 AI 可以设计出真正创新的功能性蛋白质,而不是仅仅停留在对天然序列的微小调整。24% 的成功率在蛋白质设计领域是一个相当亮眼的数字。
第二,它为基因编辑工具的开发开辟了新路径。TnpB 的小尺寸使其非常适合通过腺相关病毒(AAV)等载体进行体内递送,而 AI 优化后活性的大幅提升,使其有望成为下一代基因治疗的候选工具。
第三,这种方法具有普适性。研究团队开发的“进化信息掩码策略”不依赖于已有的实验结构,只需进化信息即可确定关键残基。这意味着它可以应用于其他类型的 RNA 引导的核酸酶,甚至是其他多结构域蛋白质。
这项研究也完美地诠释了“站在巨人的肩膀上”,这里的“巨人”,即是亿万年来的自然进化,也是近年来飞速发展的 AI 技术——自然亿万年来进化出来的 TnpB 分子,在 AI 的帮助下,几天之内就设计出了性能更优的变体。
https://www.science.org/doi/10.1126/science.aed6123









点在看,传递你的品味





